
Clades and Cladograms: Helpful Concepts to Understanding Phylogeny and the Hereditary Links Between Plant Groups
Cladistics is a system of classification…of course it is…that relates species to one another based on heredity and lines of inheritance. Before you dismiss this as a totally boring topic, consider that these are tools, that with a little study, can go a long way in clearing up the murky waters of taxonomy and systematic botany. Cladograms are a diagrammatic, graphic devices to visually display the relationships of closely related organisms, like the one below, and can be helpful to us in our efforts to understand the phylogeny and evolution of plants. The ‘branches’ of a Cladogram represent clades, all of the descendants are of a chosen ‘root’ species. Each clade must be monophyletic, complete, including all of the descendants of the root or ‘stem’ ancestor. The APG demands precision.
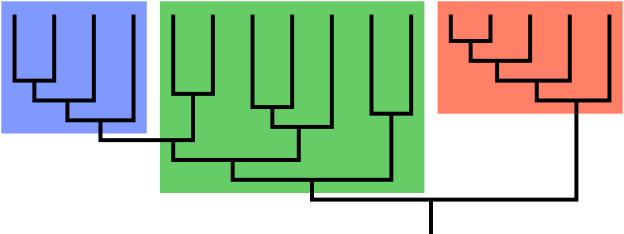
“Cladogram (family tree) of a biological group, showing the last common ancestor of the composite tree, which is the vertical line ‘trunk’ (stem) at the bottom, with all descendant branches shown above. The blue and red subgroups (at left and right) are clades, or monophyletic (complete) groups; each shows its common ancestor ‘stem’ at the bottom of the subgroup ‘branch’. The green subgroup is not a clade; it is a paraphyletic group, which is incomplete here because it excludes the blue branch even though it is also descended from the common ancestor stem at the bottom of the green branch. The green subgroup together with the blue one forms a clade again.” (emphasis mine) This is from Wikipedia, generally a good source for an overview.
Angiosperms – What is a Flower?
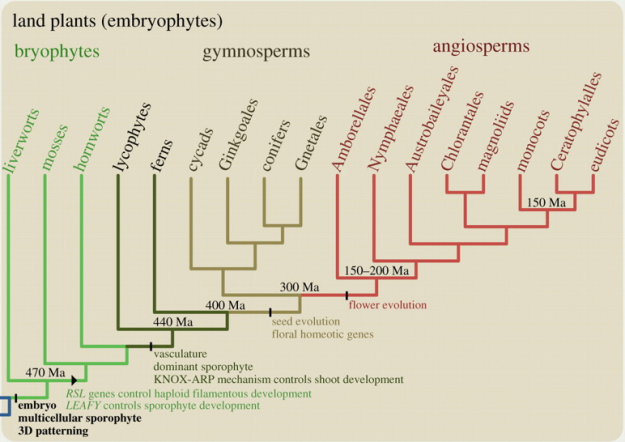
Flowering plants changed the earth. They comprise approximately 300,000 species while Gymnosperms include a total of only 1,000 today, the Ferns, including horsetails and any other vascular, non-seed producing plants, number over 10,000 species and Bryophytes, non-seed producing and non-vascular plants, have about 20,000…. Angiosperms seem particularly well adapted to the world today comprising the bulk of most of earth’s landscapes…of these, at least 70% of extant species are Eudicots, all of this occurring since 150 to 190 MYA. Angiosperms represent an unprecedented explosion in species diversity. While they possess other internal differences it is thought that their ‘flowers’ gave them their competitive advantage allowing them to diversify and speciate into forms that are so well adapted today.
Because we are looking at the ‘flowering plants’ , or Angiosperms, it is important to look more deeply into what defines a ‘flower’. It is not enough to assume that we all know what a flower is. Flowers are the site of sexual reproduction in Angiosperms. Gymnosperms and ferns also reproduce sexually and are capable of mixing the genetic material of the two different parent plants, but those do not have ‘flowers’ and this makes their process somewhat different as well. Also, many people are likely to dismiss the flowers of many species as the genuine article because they lack ‘showy’ structure or petals that they have come to expect with ‘flowers’.
For flowering plants their often showy petalled structures serve primarily to attract, guide and/or limit pollinators. Petals are neither necessary for a plant to be considered an Angiosperm, nor, technically, for the process of sexual reproduction. In many cases a flower’s corolla, and often the underlying calyx formed by the sepals, limit pollination to relatively few pollinators, by limiting their access. In some cases insects tear through the corolla to access a flower’s nectar, bypassing the pollen and anthers. Successful pollinators enter the flower in a way compatible with the flower’s structures.
Flowers possess specialized structures, that are essential to successful reproduction and are defining for Angiosperms. Stamen produce pollen, which contain the ‘male’ gamete, or sperm, within the anther on the end of a filament. Each of these anthers is divided in two, by a line that splits, dehiscing, releasing the ripened pollen which contain their haploid gametes. For each species this process continues for a specific period, which is generally considerably less than the length of time the flower persists on the plant…though many species flowers may persist for less than a day. It is not the ‘Day Lilies’ alone that only last for a day. Anthers tend to occur in multiples arranged around the pistil, the female part of the flower (some species have flowers that have separated these two structures from each other producing separate sexed flowers, in some cases producing individual plants with only flowers of one sex or the other. Both of these are more primitive traits.) The pistil consist of a stigma held above on its style which connects it to a carpel, below, where the ovule, the female egg or gamete, is produced. The ovule contains the female genetic material that will develop into the seed. The carpels in Angiosperms are always fused together forming an ovary.
Pollen is released, and is ‘delivered’ to the stigma, pollination, enabling the process of fertilization. It is important to note here that all Angiosperms under go ‘double fertilization’ as I discussed in an earlier post, one gamete fertilizing the embryo and the other fertilizing a pair of polar nuclei within the ovule that will go on and form the two ‘seed leaves’ or endosperm which will serve as a source of nutrient for the growing embryo after germination. The fertilized ovule remains in the ovary developing into a vital mature seed, an embryo and nutritive endosperm, both protected within a seed coat surrounded by the ovary which develops into a ‘fruit’. The fruit may be a simple dry husk or fleshy. The ‘fruit’ has evolved to serve a dispersal function whether it makes the seed more palatable to animals who will carry it away from the mother plant until it has been ‘released’ by the animal defecating, possess external structures that will catch in an animal’s coat or forms a structure such as the dry little ‘parachute’ that allows seed of many plants to be carried by the wind. Biologically, the tomato we eat, the husk or chaff of the wheat seed and the little seed floating on the breeze below its pappus, are all fruits, that contain seed.


The seed of Gymnosperms have no protective cover, no ‘fruit’, nor are their carpels fused into an ovary, theirs remain separate. Gymnosperms also don’t undergo double fertilization. In them the pollen grain enters the ovule and begins to develop sperm there which are later released. Emphasis on later. This takes some time compared to Angiosperms and is thought to do so because their pollen wall, exine, has no furrows or pores so it takes longer for the sperm to emerge. In some of these, like Ginkgos and Cycads, the sperm are motile, have a flagella, and swim to the embryo which they fertilize, a more primitive hold over from earlier plants like Ferns and Bryophytes. In conifers the sperm don’t have flagella. Instead they, in a way similar to Angiosperms, grow a pollen tube through which the sperm arrives to fertilize the embryo. Each seed contains more than one embryo, but only one develops as a seed, the other develops into a nutrient supply for the seed. There isn’t a separate fertilization event that forms an endosperm. Quibbling differences you might say but differences they are.
Another notable difference with Gymnosperms is that their seed is formed without a surrounding fruit. Now this used to cause me a great deal of consternation because, I thought, if they don’t form fruit, then what is a Juniper berry that we make Gin from? What is the horribly smelly fleshy coat covering a Gingko seed? and, what about the Yews with their cup shaped fruit like structures? Apparently these are other examples of convergent evolution, where these tissues formed independently from their progenitor species. Their ‘fruits’ developed from surrounding leafy bracts which fused and swelled around the seed, likely because it gave them an advantage in seed dispersal. Some of these I know are eaten by wildlife, but something at one time ate those stinky Gingko ‘fruits’? Throughout the plant world comparable structures have evolved without any genetic connection to meet comparable environmental conditions/pressures.
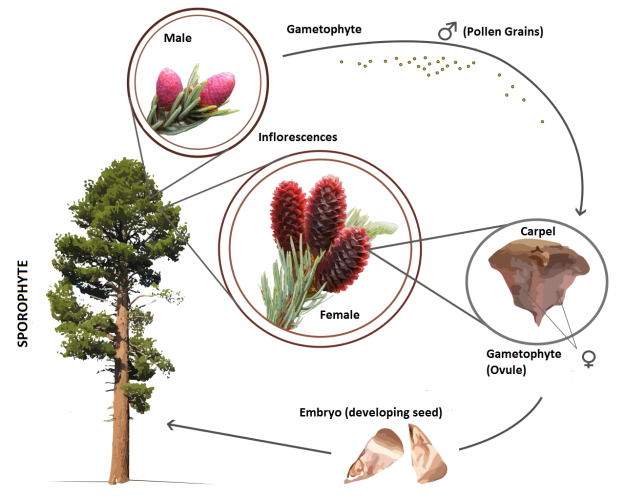
The non-flowering conifers within the Gymnosperms From Wikipedia
In Gymnosperms there are no petals or sepals surrounding their reproductive structures. There is no ovary. Seeds develop ‘naked’, uncovered. They produce pollen and ovules on two separate ‘structures’, male and female strobili. These are modified from stem and leaf tissues forming a central axis, a ‘stem’, upon which the pollen or ovules are held in a dense spiraling aggregate. The ovules are each held separately in these strobili beneath a shielding bract. On conifers and cycads, the two largest groups of Gymnosperms, the female structures develop into what we call cones, each bract is modified woody stem tissue beneath which forms the seed. In other groups the ovules form beneath small leafy bracts. On Ginkgos, there is a much reduced cone structure the ovules forming on short stems toward the branch ends. The male structures are shed after releasing their pollen. (See my previous blog posting for a more detailed explanation of the different forms of sexual reproduction in plants.)
Angiosperms and their flowers are more complex in their structure and function than the strobili of Gymnosperms. In flowers the process is more efficient and direct. More ‘energy’ is invested into each successful seed and endosperm and their evolved methods of seed dispersal. In some cases they have also developed the ability to ‘self’ increasing the likelihood that even small isolated populations can increase their numbers…The more recent fossil record also helps underscore the more recent trend in Angiosperms of inflorescences with compound flowers that through their density increase the ease of pollination. The modern Asterids are a good example.
Many thousands of species are extinct and left behind no fossils. These extinct species would have been intermediate steps along the development path. Angiosperms, most scientists agree, first appeared on the scene around 139-125 million years ago, + or – a few million. Part of the difficulty in pinning this date down more solidly is that there were several ancient groups of seed plants within the Gymnosperms and their predecessors, that had characteristics that were closely related, morphologically and genetically to modern flowering plants. Again, it was not that they didn’t exist one second and did the next. In a sense there is a threshold for ‘Angiospermness’, how many of the traits are necessary in order for us to label a plant as an Angiosperm(?) in what seems to be a very incremental process. It is difficult to agree when precisely they first crossed that threshold. Since their arrival, however, Angiosperms, have been extremely successful.
On the other hand, the Gymnosperms, which for many millions of years, dominated most of the Earth’s regional flora, have long been in decline. Their arrival and success contributed to the changes that tipped the conditions to favor Angiosperms. Angiosperms were more prolific given their reproductive improvements, with many of their species serving as early successional weedy type plants, over time diversifying adding species that could form canopy over-story trees in diverse rain forest type landscapes…that, in many landscapes could out compete the older existing Gymnosperms of the time.
TheExtinction Events and The Rise of Angiosperms
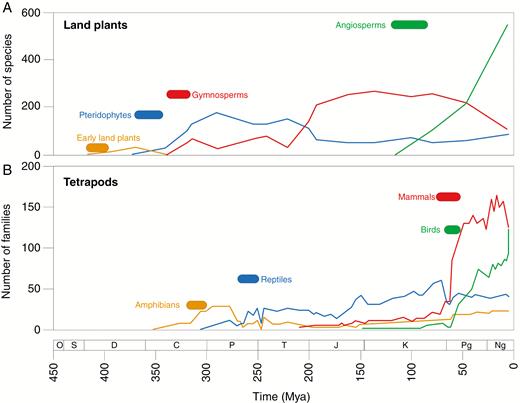
This chart shows the major plant and animal groups over the last 400+ million years, mammals and angiosperms being the new kids on the block. The Cretaceous-Palaeogene extinction event, about 66 million years ago, can be easily seen on the, lower, Tetrapod (four legged animals) chart. There is a distinct drop in Reptilian families and the other groups as well, followed by a rapid increase in mammal and bird families. There is no corresponding drop and rise amongst plants indicated on the upper chart. The earlier extinction events also show pronounced drops in animal families.
Many argue that the big extinction events that have occurred sporadically over history, in a sense, cleared the board of many of the more ancient species providing niches and opportunities for the ‘newer’ species. This resulted in the two-fold process of speciation and radiation as these new plants spread and adapted…but, most of these extinction studies focused on animals and the record is much more ambiguous regarding plants. Most pertinent to Angiosperms is the Cretaceous-Palaeogene (CP) extinction event about 66 million years ago, the most studied of the ‘Big Five’ events due to our interest in Dinosaurs. The CP extinction was the result of several ‘events’ coinciding closely with one another including the asteroid that crashed into Earth at the Yucatan peninsula, a large increase in volcanic activity and the consequent release of massive amounts of sulfur compounds along with global sea-level changes. This is thought to have resulted in the loss of over 80% of all marine invertebrates and a drastic reduction in mammalian species including entire genera and families. For plants the impact was less devastating losing individual species while genera and families suffered reductions in numbers, but rebounded back in ways responsive to their changed conditions. Animals, it would seem, live within a narrower range of parameters. The changes that did occur in regional floras are thought to be brought on by changes in climate largely due to the impact event..
Across the globe, leading up to the CP extinction, landscapes were already dominated by Angiosperms. This was a time of incredible speciation and radiation, their numbers increasing rapidly. It was an event defined more by bio-mass reduction than it was species extinctions…at least for plants. Growing conditions were significantly changed by these events tipping the balance, favoring one species over another, but not to an exclusion. In many cases deciduous plants had a significant advantage and came to dominate many of the previously dominant evergreen species, pushing them into smaller niche landscapes, not extinction. It is thought that no families within the Angiosperms were lost during this period. All survived. Even those lost from particular regions are thought to have later recovered, their seed slowly moving back into landscapes as their climates returned to something more similar to pre-event conditions, though this may have taken a million or more years. Some clades benefited greatly during this period, increasing their numbers and spread well beyond where they had existed previously, occupying landscapes that may have earlier been dominated by taller growing Gymnosperms. The balance between species changed with those less competitive consigned more to the margins.
This event, as had previous ‘extinction’ events, resulted in an immediate reduction in numbers of woody plants worldwide while herbaceous plants and spore producers, briefly became dominant, only to be supplanted later by woody plants, changing again those that would dominate. The new dominate plants were present before, but had gained a competitive advantage during the event. Woody Angiosperms took their dominant position across the majority of forested and shrubby landscapes.
Today there are arguably more different species alive than at anytime in Earth’s history, though it is difficult to support this definitively because so much of the past is not preserved in the fossil record. Extinction events have consistently been followed by ‘explosions’ in speciation and radiation, ‘innovations’, followed by many millions of years long periods of consolidation and relative stasis, before the next such event. Angiosperms, and the Eudicots, are still in the ascendant position. What comes next, and when that may occur, is something well beyond anyone to say…but such a ‘cycle’ is more than likely to come around again. But, for now, and in a foreseeable future that could last many more millions of years, Angiosperms, and the leading edge of Eudicots, will remain in the forefront…barring some incredibly tragic act of hubris on our part like the disruption of the Earth’s natural systems to the degree that they can no longer continue in a way supportive of life….Historically, in a sense, the loss of species has always been a result of the further development of the Earth’s living, natural systems. A building toward greater and richer complexity, not simple consumption into a downward spiral of entropy and dissolution…though this was not without its losses and sacrifices.

Pingback: Kingdom Plantae | Taxonomy 3 – ChukarBlog
Wow Lance! What a tremendous amount of research and writing you’ve done here. I better start my reading . . .
LikeLike