
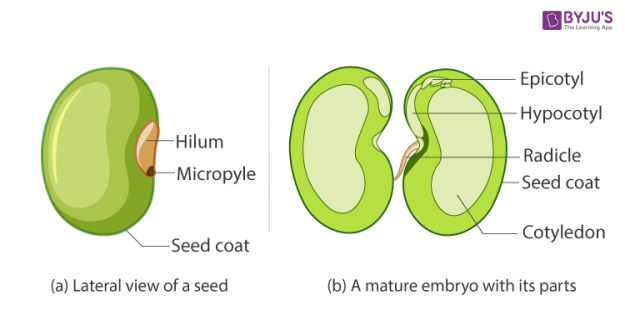
This diagram is of a typical bean seed here to illustrate some of the basic structures within all seeds. When germination begins water is taken in via the Micropyle. This picture is taken from a simple introductory page, https://byjus.com/biology/identification-of-the-different-parts-of-an-embryo-of-a-dicot-seed/
EVERY, let me say that again, every, single individual organism, plant, animal, fungi and bacteria…is directly linked, in an unbroken line, to previous organisms. An organism is not created singularly and anew within a Frankensteinian lab, whether of our own hand or nature’s. Life is an extremely rare occurrence. Genesis did not happen, in the biblical sense, but on extremely rare occasions, arguably only once in Earth’s several billion years. The conditions it requires are unique, precise and stable. Just as individuals are linked directly to their parents, entire species are to their predecessors. So called ‘spontaneous generation’ does not happen. The idea that organic matter can be manipulated and ‘sparked’ into life is naive. It can only be more or less manipulaed as we’ve demonstrated time and again in a heavy handed way. Science, especially over the last one hundred years, has made great strides in understanding just what life is, what it requires and how it most likely evolved, but it still cannot ‘create’ it. Even in its most simple forms, such as bacteria, life requires the ability to conduct thousands of biochemical processes within each cell in a very precise way, something that not even a series of highly coordinated human operated laboratories, using standard of the art equipment, can do in anywhere near the amounts and efficiencies that a single living cell can. We are far too clumsy. Doing this for a complex multicellular organism with highly specialized cells, tissues and organs would seem impossible. As we continue to study organisms, their processes and growth, we are learning just how complex and astounding they are.
When we propagate individual organisms we do not create life we only permit a natural process to move ahead. We choose one individual over others. All organisms contain within themselves the capacity and drive to reproduce. Successful reproduction, for many species includes the rearing of one’s offspring to maturity through nursing, feeding and defending. Reproduction is linked to an organism’s lifespan. Many plant species are being found to share direct links, through their root systems, or via mycelial networks, whereby the can share resources and signal one another providing a competitive advantage to juveniles and those in need. Plants thus grow in ‘communities’, members both in competition with others and supported by still others. Over several centuries we have refined our ability to manipulate this capacity to serve what we see as our own humanly defined interest. These links and relationships have often been overlooked, while at the same time we have a strong tendency to see each individual as entirely separate organisms. This oversight has resulted in our weakening of natural communities. Hopefully we will begin to recognize the role of these many relationships for the health of the whole. As we continue to disrupt the landscape and such communities on a global scale, this has become of much greater importance. We have long taken advantage of the capacities of other organisms to reproduce, grow and heal themselves, leaving the health of natural communities to dwindling sites. Life is a ‘package’ deal. Our cleverness in propagation is only part of the picture and we need to understand that we haven’t created new life when we do so, we have only utilized what is given.
All organisms, which reproduce sexually, undergo remarkably similar stages. First, the single chromosome gametes are formed; Second, the male gamete, the sperm/pollen, fertilizes the female gamete, the ovum, forming the zygote; Third, the zygote develops into an embryo; Fourth, over time, and under the right conditions, the embryo develops into an individual capable of independent living. Upon reaching maturity this individual is then capable in participating in the reproductive process. Each species follows its own particular path while individuals of the same genera and families may share much in common. The different species are linked directly following their genetic lineages. Whether reproduced sexually or cloned, they share inheritable traits passed from one to the other. Central to this they share that spark of life, ‘handing’ it off to the next generation, or as a clone, the next iteration of itself.
DNA is often presented as the ‘magical’ and managing component within living organisms, but it is not the energy of life, nor is it the structure that defines it, a blueprint for a self-constructing organism…it is an incredibly complex set of instructions for operation and maintenance. Mass is sometimes defined as ‘informed’ energy, energy that is structured in such a way to produce a substance’s recognizable characteristics. ‘Informed’ differently changes the mass. When broken down into quanta, those unimaginably tiny bits that cannot be broken down any further, matter would appear to have little substance at all. Living organisms are composed physically of the same stuff that comprises other matter, only the ‘recipe’ would appear to be very different in its composition and along with it, its associated energies, energies that are not found in inert matter. Whatever the species or its maturity, living organisms are animated by a particular energy that separates it from a simple mass of organic compounds, cells and tissues. Could we ‘build’ a steer from grass, oats and hay, it still wouldn’t walk or moo, just as we cannot draw carbon and nitrogen from the air, minerals from the soil with water to grow a plant without a seed or bits of pre-existing living material. Our ability to ‘create’ life is limited to beginning with living organisms and modifying them.
In vitro growth of an organism still requires a living embryo, or in specialized cases, actively dividing stem cells, that are, actively growing and dividing. An embryo is the product of an ovum and sperm cell, two oppositely sexed gametes, each of them ‘alive’, but not sufficient on their own, neither able to grow into a mature organism. That pollen grain, depending on the species, can lie around dormant, under the right conditions, for many decades, sometimes much, much longer…still ‘alive’. In a few species, such as the ubiquitous Dandelion, a plant can be capable of apomixis, a process in which the unpollinated flower, the unfertilized ovum, can still produce a viable seed, but this is uncommon. In general, a viable seed requires fertilization. Many plant species produce seeds that can remain dormant for centuries, which can later be germinated….Seeds exist in a state of suspension, their metabolisms switched off until dormancy is broken, imbibing water, after experiencing a ‘cold’ or hot period, the acid conditions of some organism’s digestive system, the physical scarification of the seed’s seed coat, fire and smoke or some combination of these. While it is dormant, a seed is poised on the edge of life. Once germination begins all of the necessary steps in life follow in a ‘stream’, continuously, if life is to continue…it must. Every organism then lives within its metabolic limits without ceasing.




Seed dormancy varies and is specific to each species maybe even to particular populations from a given region. Determining what is required to break dormancy is essential to get the best germination rates for a seed propagator. This makes perfect sense when we understand that all plants follow very particular patterns, initiating growth each year, following a developmental ‘calendar’ through the year, flowering, fruiting, dying or returning to a state of dormancy induced by cyclic and seasonal changes. These conditions will reflect those historically in effect across a species native range. Random germination throughout the year, responding to quick changes in weather, for example, would result in a much higher failure rate, reducing a species chances of maintaining a viable population. Such patterns increase the chances that germination will result in establishment and growth. Temperature requirements tend to prevent germination until the time of year when establishment is most probable. Requiring a sustained cold period helps prevent premature germination in the Fall which could result in the freezing deaths of seedlings over winter. Having a fire/smoke requirement is likely resultant from the advantage of fire clearing away competition from less competitive seedlings. Other species require acid treatment as a result of successful seedlings carried further from the ‘mother’ plant in the guts of animals who’ve consumed it. Others require light perhaps delaying the germination of seeds buried too deep, which would otherwise cause them to expend the limited energy contained in their endosperm before they can reach the surface, before their leaves can emerge and capture the energy of the sun. Seeds of species from the wet/tropics will have very different requirements than those from high cold steppe, chaparral, alpine, tundra or temperate prairie regions. Whichever seeds we intend to germinate we need to understand this.
The germination of any one seed is regulated by a complex array and balance of phytohormones produced by the mother plant and placed within the embryo at its formation where it resides within the seed, which later work with the environmental factors to allow germination under the most favorable conditions. These biochemicals like gibberellic and abscisic acids, auxins and other compounds work to variously impede and release germination. Together the keep the seed in dynamic tension, on the verge of growth…ready. Positive, supportive conditions lead to the oxidation and breakdown of impeding compounds allowing those supportive of it to prevail and ‘slingshot’ the embryo into growth.
 The seed embryo is developed as the seed forms, in flowering plants while it remains within the flower’s ovary. This process of ’embryogenesis’ follows dependable patterns of cell division and differentiation…it doesn’t just suddenly appear in its entirety. For those interested, check this site out. The above diagram is from it.
The seed embryo is developed as the seed forms, in flowering plants while it remains within the flower’s ovary. This process of ’embryogenesis’ follows dependable patterns of cell division and differentiation…it doesn’t just suddenly appear in its entirety. For those interested, check this site out. The above diagram is from it.
One of the first changes within the seed at the onset of germination, is its imbibing, taking on, of water and O2 from outside via the micropyle, a limited tiny entrance to the seed interior. The O2 enables the needed oxidation of inhibitors, a common chemical reaction process occurring everywhere within cells, a natural process that the cell, the organism must continually offset to remain alive. Water is itself a chemically activating agent (see Gerald Pollack’s work in, “The Fourth Phase of Water”.) playing an important electrical and structural role within the cell and organism, perhaps setting the stage for much of its function. Chemical bonds are electrical in nature and water, as a strongly dipolar molecule, possessing both positive and negative poles, attributes unique and essential properties in its fourth state, to produce charged hydrogen ions, central to many of the cell’s processes.
Once released through germination the architecture and micro-architecture of every organism, shaped with such specificity, inform the development and operation of each individual, the bio-chemical dominos tipping in concerted action, every protein and enzyme poised and ready, coordinated, in synch, and inevitable. The engines of the thousands of mitochondria and ribosome, its other organelles, in each cell are primed and ready to spin cranking out proteins, enzymes, all of its metabolites, on an as needed basis, including the biochemical batteries, the ATP, which powers the countless chemical reactions necessary for every living cell every second. Continuous throughput of energy, nutrients, water and electrolytes are required in proportion to an organism’s size, complexity and rate of metabolism. This is the classic chicken or egg problem and the answer is still both, one does not exist without the other.

Seeds are incredible structures occurring in sizes that range from that of dust to that of the Palm pictured here, Lodoicea maldivica, the Coco de Mer, a rarity limited to the Seychelles in the Indian Ocean, that forms the seed above that can grow up to 12” long and up to 40lbs within a truly massive fruit that can push the total weight to over 60lbs, taking 6-7 years to develop! This is a dioecious species with separate male and female plants, the males of which produce a meter long catkin like inflorescence which persists and can produce pollen for as long as 10 years! It was once known under the specific epithet, callipyge, from the Greek words meaning ‘beautiful buttocks’…I’ve always liked the more descriptive names rather than those linking a species to a botanically significant person. With that massive endosperm much of the emerging seedling is large as well. A tube forms between the seed and developing seedling that can be over 15’ long. The emerging cotyledon, its ‘seed leaf’, is the largest in the world at up to 14′ long! Its large endosperm contributes to its ability to withstand stresses during establishment. Seeds and seedlings have highly variable seed coats and dormancy, effecting their ability to maintain their integrity while withstanding the conditions it may be exposed to. Seeds of tropical plants such as this tend not to require cold stratification, surprise, surprise, their key often be associated with moisture due to the two seasons, wet or dry, that the tropics experience. Generally, in this species, each seed contains a single embryo. Contrary to this there is little margin for error with tiny seeds, such as those for Begonia, with their proportionally tiny endosperm. As with most seeds the bulk of the Coco de Mer seed is endosperm…but, as in any seed, it is the embryo that contains the ‘magic’, the spark of life that will ignite into the seedling.
Seeds should be viewed more as a means by which life is passed from one generation to another than they are beginnings of distinct independent living organisms…there is no break or true beginning. Each individual is an extension of a line…a continuation. While the individual dies, the species, the line, continues and that is what is important.
As gardeners and horticulturists we generally select the plants that we grow, disrupting the natural selection process, altering the composition of the communities that would normally arise, while the weeds seed in and find their own positions. Weeds, through the genetic mixing of sexual reproduction of successful individuals, are often very well adapted to the conditions on a site and contain a broader range of genes, along with their adaptability and ‘toughness’, to survive and increase. By ‘choosing’ we narrow the genetic range of what we plant. Even when we grow our own seedlings, that seed is somewhat limited genetically by our choosing as we collect and share seed from a particular plant to friends and fellow gardeners. Sometimes commercial seed companies do the selection from particular ‘strains’ they market, sometimes hybrids.
Clones and Vegetative Propagation
Many of our plants are clones, divisions, rooted cuttings, grafted or grown in a lab utilizing the several techniques of tissue culture. From a single plant, exhibiting particular characteristics, grown from somatic, vegetative tissue, can come thousands of identical individuals, over relatively short periods when tissue culture is successful, much less when grafting or other more traditional methods are used. Somatic plant cells, though they can be specialized themselves, all contain a nucleus which hold the plant’s entire DNA, which is later ‘expressed’ in the mature, cloned individual. Most of a plant’s tissues are composed of somatic cells, those not involved in the production of single chromosome containing gametes. An embryo is somatic. If somatic tissue has the capacity to develop into a viable plant, and not all of it does, the life it contains can be used to ‘jump start’ a ‘new’ plant, an extension of the former.
In plants it is generally younger stem, root, bud or leaf tissue that is used in propagation because these contain actively dividing meristematic tissue, active sites of cell growth and differentiation. Much living tissue does not contain these ‘stem’ cells and cannot be successfully propagate or does slow only with difficulty and much lower rate of success. Stem and root cuttings are ‘sheathed’ within actively dividing meristematic tissue, its cambium while buds contain it which is capable of continuing the top growth of the plant or, in other cases, more specifically, its flowers.
Once cut off and separated from the ‘mother’ plant one must proceed quickly to assure that these tissues stay viable. Their must be little break in the flow of water and nutrients to these tissues. Removed from the ‘mother’ plant they must be kept hydrated and cool to slow metabolism. In the case of grafting, this work is generally done during dormancy, just before the onset of spring growth. Cuttings can be taken and rooted across much more of the calendar with greater or lesser degrees of success. Often, their meristematic tissues can be stimulated into more active cell division by utilizing plant growth hormones in specific concentrations, forcing them into forming callous tissue at the wound site from which later roots will develop. Generally, top-growth, stem and leaves, will form from the still healthy and included bud tissues on the cutting or graft. Whichever technique is utilized the cut tissues must be protected from desiccation by placing it in a relatively moist environment while also guarding against fungal infections…a priority must be placed on sanitation.
Leaf tissue is less commonly successful, but in some genera and species, remembering that expanding leaves arises from a self-limiting and thus ‘determinate’, meristem at its margins, leaves can be sliced transectionally, perpendicular to the leaf’s central vein, and placed in a moist rooting medium on edge to develop tiny plantlets.
From life comes life. Plants have relatively slow metabolism rates so the exposure and injury that clonal propagation techniques imposes on them is not immediately fatal. They have evolved defensive strategies to contain or limit wound damage very different than animals. Bottom heat systems are often utilized to warm and speed root growth at the base, while the tops are kept cooler to slow cell metabolism until roots that provide for it are formed. Successful procedures are often quite specific. For many they require a ‘kickstarter’ often in the form of the application of plant growth hormones in appropriate concentrations. Time of year and the condition of the selected tissue are important. What works in one case won’t in another. In other genera and species the same techniques will yield relatively poor results. Propagators build their careers around the specialized knowledge of particular techniques applied to particular tissues at the appropriate time under precise conditions. Low success rates can ‘kill’ a business.
Nurseries ‘protect’ and market their clones intensively, every plant sharing identical genetics. To the extent that such clones succeed, the gene pool shrinks and the species survival can become problematic as gardeners and growers choose it over seed grown stock. Often, especially when numbers of wild plants are very low or long distances away, cloning may be the best or only option to increase numbers. We should remain aware that, in certain situations, cloning can compound the problem. Gardeners who, with our aesthetic biases, tend to ignore or neglect out of hand, many thousands of species. The problem is compounded as intact natural populations decrease with shrinking habitat. While cloned plants are as ‘alive’ as seed grown, their ‘market’ success can lead to a less stable situation for the species. Garden grown plants may be healthy, while the state of the species can be precarious. Garden grown plants are dependent upon the gardener into the future as native, intact landscapes diminish reducing the native populations.
This is the situation for an increasing number of species such as the Chocolate Cosmos, Cosmos atrosanguineus, which is extinct across its natural range in Mexico and Arctostaphylos franciscana, the last surviving, wild, individual found on a neglected site in San Francisco’s Presidio in 2010, which itself was nearly lost to road construction. Today it has been successfully cloned and transplanted, sharing quarters with a handful of clones from a few other surviving plants found living for decades in private gardens. These have all been planted on a protected site in hope that a natural community can establish, in spite of the fact that its genetic material is drastically reduced, the rest all lost to the development of the city. Traditional garden practices, working with urbanization and the spread of agriculture, combine to reduce natural plant communities and prevent the establishment of new ones that could assure the survival of a species into the future.

This picture is a section of a Gingko biloba seed with its embryo attached to the endosperm via to supply it with nutrients at germination, taken from Wikipedia.
While all tissues and cells in animals are alive, metabolizing compounds, replacing proteins, maintaining the components of their structures, this is less true of plants particularly woody Eudicots and Gymnosperms, which contain ‘wood’, inactive, static structures which support the living tissues which encase it. The ‘softer’ tissues of plants are still very much alive, but they tend to grow in a determinate, limited manner, though some can grow for centuries into very large structures, they remain recognizable in their parts. Plants don’t grow into amorphous blobs, random shapes or with branches, leaves and flowers of unlimited size or shape….A leaf or flower grows only so large, in a very particular shape, before it stops and is eventually shed and later replaced. Stems and branches follow patterns as well determined by their genetics and the conditions under which they grow. The organism, the individual is alive, functioning as a vehicle for the genetic line, ‘expressing’ it. The individual lives for the ‘line’. We might celebrate particular individuals, but other organisms don’t. They simply succeed or don’t and the determiner of that success is the ‘health’ and vigor of the community. The criteria are dynamic and changing.
Plants do not require us, but we do them in complex ways, in part because we as heterotrophs must consume them to sustain ourselves and because the health of the larger community, of which we are a part, require them. We have a role to play in this community in order to sustain it, a role we have largely ignored, but one who’s consequences loom ever larger as our negative impacts increase. Our ‘success’ is contingent upon how well we understand them and work to meet their needs that other species no longer can, due to our disruptions, by how well we can ameliorate and compensate for the ‘damage’ we have done. The further we take them from their natural conditions, the living communities within which they evolved and live, the more we must do to meet their specific needs, and we are doing this poorly. When we don’t, their health begins to fail and we find ourselves in the role of ‘nurse’. Life is ultimately not separable. The independent individual is a myth. All living things exist in dependent relationships…a fact we have been ignoring and dismissing for most of our time here as a species.
Life surrounds us! Its ubiquitousness lulls us into believing that it will go on ‘forever’…and it likely will, just not in the form that we might expect. Each species, each population, exists as a lineage, in a continuous line, that once broken, ceases to exist along with all of its possibilities. We cannot create it anew. We are limited to the role of steward, husbanding the life that is already here. When a strand fails, the whole is altered forever. The last Black Rhino, once lost, is simply, and unequivocally, gone. The same is true for a rare Orchid or any other species. Movies like Jurassic Park, promote the idea that life can be recreated from samples of intact DNA, but DNA is a ‘code’ not life itself. Life exists in a package. It cannot be stored in a computer or archived to be replicated on demand. While climate controlled ‘seed banks’ are used to store seed and thus protect their germ plasm and the genetic diversity it contains, that seed must still be occasionally tested and grown out to replenish the stored seed. Life is perishable. If we truly want to assure the continuation of life on this planet we must protect intact and viable landscapes around the world so that the germ plasm can be ‘protected’ in the form of living, still evolving, plants, in much the same way the a landscape, like the Amazonian Rain Forest, is best preserved by conserving its many communities as living, healthy systems and landscapes. That is where life springs from.
While biochemists will argue that the chemical reactions of life are ultimately electrical, the Frankensteinian notion that inert organic matter can be sparked into life, is naive. Life exists in the moment in a continuous chain of perfect actions and reactions. Decay begins immediately at death. Life requires a continuous flow of water, nutrients and energy. The life within the first organism continues today in the form of billions of living organisms, trillions if we include bacteria. The idea that we can create it fresh is an arrogance. While we may describe an organism in mechanistic terms, life is not built from component parts. We may fiddle with it, tweak it here and there, but if such efforts stray too far our experiment will fail, quickly or some time later. After all, death is an integral part of life. The continuation of the species requires it. Organisms exist in the dynamic space between the two. At its heart life is the root of all that is sacred. The sacred is not separable from life.
Interested in the reproductive cycle to see what precedes all of this? This is a great explanative diagram. It come from the same site on which I found the one for the development of a plant embryo.
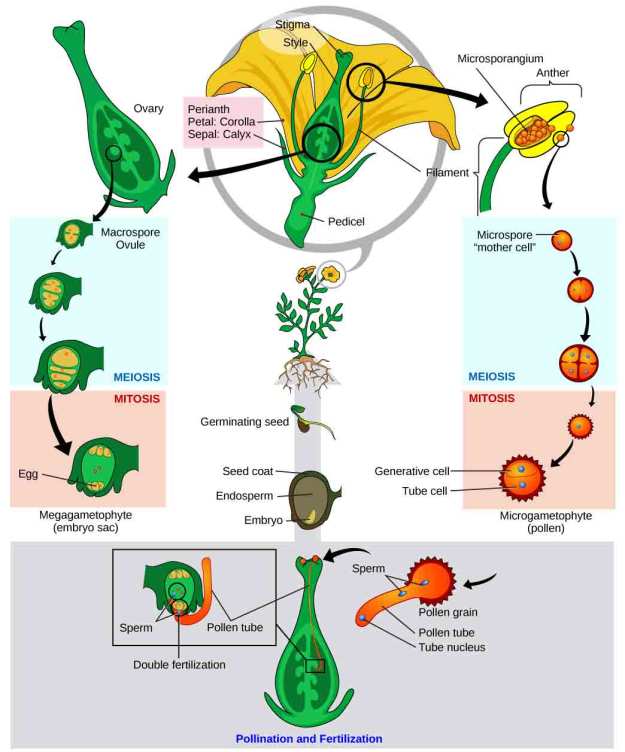
This was found on the site, http://www.yourarticlelibrary.com/biology/embryo-in-flowering-plants-structure-types-and-development/11808
