I use a lot of Monocots in my garden, among them in this picture are the Colocasia esculenta ‘Black Magic’ with its huge dark velvety heart shaped leaves, Arundo donax ‘Variegata’, the Giant Reed, whose clasping leaves show us that this is a grass not a bamboo and the white speckled, green, heart shaped leaves of my Zantedeschia x elliotiana ‘Flame’ just behind the reaching stem of Arundo.
Many gardeners are self taught and haven’t formally learned Botany, the science that helps us understand plants in a more formal, academic way, though they may be excellent ‘gardeners’ in terms of their growing of plants. Botany provides a pathway toward the understanding that many of us crave, that for others is an unwanted burden..they are happy with the doing. For them the task of learning botanical latin, binomial nomenclature and the classification system by which we organize and study the various species, understand their structure, development and common history…is of less interest. No doubt a good many fall somewhere in the middle. I have always been among the more curious ones with regards to this.
Fitting Our Garden Plants into the Plant ‘Universe’
At 30, I went back to school to learn horticulture and enough Botany to give me a formal framework to understand what I had chosen to do professionally. Without this I was struggling. Every plant was an ‘individual’ and as I learned more plants it was becoming harder to keep them in my head. I craved ‘context’ and a way to organize them…for my own benefit and so I could better understand others when they spoke or wrote of them. What I found was initially a bit overwhelming, because not only were plants organized by genus/species they were related more generally within families, orders, divisions and on. Plant-people would talk about various plants within these groupings and it wasn’t a random arrangement. They were related with overlapping and branching relationships, shared ancestry. Related plants had shared ‘morphology’, outward physical characteristics and internal anatomy, through which they could be identified and of course, because that would be too easy, there were exceptions, plants that ‘looked’ like one thing superficially that turned out to be quite another, that required us to know what to look for to properly identify them, e.g., Maples without ‘maple’ leaves and other plants, like Sycamores, with ‘maple’ leaves, but alternate ‘phyllotaxy, or leaf arrangement, and entirely different flower structures. There were other more basic differences between larger groups of plants, characteristics determined many millions of years ago, that would come to shape the species we have today, passed on but still identifiable, if one is looking. These are ‘grosser’ structural differences that can go to the very shape and function of a plant such as whether a plant is vascular or non-vascular, flowering or non-flowering, even plants that don’t photosynthesize their own carbohydrates, something that most of us would have said are most basic to what a plant actually is (Plants that are parasitic or ‘semi-parasitc). So, where is all of this going right now?
Because we ‘need’ a beginning point, life began in this world as single celled organisms, prior to that there were photosynthetic bacteria, protists and other things none of us would ‘mistake’ for plants. This was before there was even an atmosphere with Oxygen. The first plants were non-vascular, they had no veins to transport fluids, nutrients or the metabolites they produced to regulate their function, and they reproduced without the intermediate step of seeds, through cell division alone, forming largely undifferentiated masses of like cells. These were followed by simple vascular plants that formed spores instead of seeds to reproduce, like the Ferns, Tree Ferns, Horsetails and Club Mosses. Later yet came the Gymnosperms, which were vascular and reproduced from seeds developed from ‘cones’, or stroboli, including the conifers, Ginkgos and Cycads amongst others. All of these predate Angiosperms, the true flowering plants, by many millions of years. While the Gymnosperms utilize pollen and produce seeds, their seeds are not formed inside the ovary of a flower…their seed is formed ‘naked’ without the cover of a carpel, often within a ‘cone’ structure, in species within the group or Division we know as Conifers, that upon ripening, open, exposing their uncovered seed directly. Ginkgo produce them at the end of short stalks. Gymnosperm seeds once developed on other structures, on now extinct species. They once dominated the landscape sharing space with the vascular, non-seed forming, Ferns . Conifers still contain many important and vital species, while the Cycads are threatened, due to climates that have changed over millions of years and the much more modern pressures of habitat destruction and ‘poaching’ by collectors.
Angiosperms, with true flowers, dominate today. They form their seed in a flower’s ovary around which a carpel and/or fruit forms. Angiosperms began developing with the appearance of pollinators and have since been very successful. There are an estimated 400,000 species of plants in the world, estimated because species are still being found, identified and described. The Grass Family, one of the members of the Angiosperms, the Flowering Plants, occurs on and dominates much of the land surface, is comprised by something less than 12,000 species. Yes, Grasses are flowering plants and are among the many that are wind pollinated, no bees, flies etc. required. Their flower development took a slightly different ‘path’ than those that evolved ‘relationships’ with pollinator species. Wind pollination is a much more ancient strategy. Regardless of how they are pollinated, their ‘flowers’ produce seeds. That seed, possesses one of two characteristic structures that is obvious upon germination. They are either Monocotyledonous or Dicotyledonous (Monocots or Dicots)….They have either, Mono, one, or Di, two, cotyledons, or ‘seed leaves’. (Gymnosperm seeds can contain much smaller, divided, cotyledons, however, they rely on a more ‘primitive’ structure to nutritionally support the embryo of the germinating seed, a remnant of the ovary it formed within. Their emerging seedlings exhibit neither of these two structures. For those of you who might want to ‘geek out’ more on Gymnosperms and their seeds, proceed to the end of my posting and follow the link to the site.) These plants continue down different paths, each carrying with them defining characteristics that they will manifest/exhibit throughout their lives. While this may sound relatively inconsequential, it has much to say about how these two groups of plants are structured, how they will grow and how we should care for them.
The Phylogenetic Tree

A phylogenetic ‘tree’ of the plant world produced by the US Botanic Garden.
The phylogenetic ‘tree’ in the above illustration, is, obviously, diagrammatic. It displays the development of plants, from their ancestral roots moving up its trunk in time out into its many branches, showing when each group began, relative to others, and ‘how’ they are related. Each ‘branch’ contains plants with specific shared characteristics. Those closest to the roots are the oldest most ‘primitive’ of plants. The four golden ellipses identify the most basic structural developments over time. The largest limb, bending and sweeping off to the right, represents the ‘Angiosperms’, the flowering plants, and divides into two, the larger of which and the youngest, being furthest from the base of the trunk, are the Dicots. Various smaller branches erupt singularly or continue on in time forming ever finer branching. The point from which each of these branches arise indicate a significant change from the plants that preceded them. Each successively smaller branch, whether it includes one Family or several share more in common than they do with a Family located on another secondary branch. The Grasses then share more with Pineapples and other members of the Bromeliad Family than they do with the Bananas, Cannas and Gingers, though these share more with each other, than they do with species of the Amaryllis Family which is on a different secondary branch, but again all of these share more together than they do with the Dicots.
’Trees’ like this have been used since Botanists became serious about the links between plants, their shared and uniqued histories. Over time the branching has changed, in the way that it’s been drawn, with our understanding and has been historically based on morphological studies and their anatomy, the study of their internal structures and outward appearance. Today the finer details are changing due to the precision of DNA studies. ‘Mapping’ the chromosomes of various plants show direct links between species that were previously not understood or recognized. Keep in mind as you look at this that each species shares many characteristics with all of the rest in the same Genus and likewise, that all of the Genera of a particular Family share many characteristics. At the top of the ‘tree’ is a small branch labelled Amaryllis. Amaryllis here represents an entire Family. Every genera and species that comprises it will be found here, there won’t be any scattered within the Onion Family or the Agave Family. All of the branches have readily identifiable characteristics that distinguish them from the others. The major limb oriented vertically, representing ‘Monocots’, is what I will be looking into here.

This pic, from 9 years ago, shows part of my outer garden from the street including several of my more substantial Monocots: Musa basjoo peaking over the fence to the left and left to right along the curb, Nolinia ‘La Siberica’, Festuca ‘Elijah Blue’, Yucca rostrata ‘Sapphire Skies’, Trachycarpus fortuneii, a blue form of Agave harvardiana and a flowering Yucca gloriosa ‘Tri-color’ with our gate framed out of solid calmed bamboo. Iris x pacifica and Kniphofia are scattered at ground level.
What is a monocot? What distinguishes it from other plants and why should it matter to you? This is a link to a fairly basic description. Turns out Monocots are much more than simply plants with a single seed leaf. There are a host of shared characteristics, some of them unexpected…some of which aren’t quite ‘universal’, but are in fact, technically true. And, like many ‘rules’, while in general true across their constituent members, there are exceptions.
Flowers and the World of Plant ‘Exceptionalism’


All Monocots are Angiosperms, flowering plants, which is why the ‘tree’ shows its branch originating from the larger Angiosperm branch. They all have true flowers that produce seed within a protective structure, a carpel or fruit. Their seeds are not formed ‘naked’ as they are in Conifers and the other Gymnosperms. On many species of Angiosperm some may argue, ‘Where’s the fruit?” These seed are formed within a flower’s ovary which becomes the fruit and makes these Angiosperms. What about Grasses and Bamboos, you might ask?…they don’t have ‘flowers’ and where’s their fruit. First, not all flowers have petals and sepals. They don’t all form a corolla. Some are ‘apetalous’, without petals, but they still possess the ‘working’ parts of a flower. The pretty flowers that we picture in our minds as ‘flowers’ have evolved over many millennia to attract pollinators, but not all species of Angiosperms require the ‘assistance’ of a pollinator. Flowers were not created for our aesthetic benefit. They developed millions of years before our species existed and relatively lately have become more obsessed with the aesthetics of flowers. Their various forms, colors and scents served to attract sometimes very specific insects, birds and bats to aid them with pollination. Flowers have only relatively recently gained a kind of human luxury status and are being grown for their own sake. Previously, indigenous people’s, while they may have selected some for decorative purposes, much like they might have feathers of birds, more likely used them as ‘markers’ or identifiers in the landscape, helping them distinguish plants one from the other for more practical purposes such as food, medicinal or other ‘economic’ uses.


Grasses and Bamboos are among those with apetalous flowers and have functioned very well with the wind distributing their pollen. Typically these flowers are small though numerous on a plant, crowded into structures on their inflorescences often in the form of spikes and panicles. They are absent scent or nectaries as they have no reason to attract pollenating insects. When looked at closely grass flowers do carry the 3 anthers characteristic of Monocots.
The cone bearing Gymnosperms utilize the ‘path’ of wind pollination, as well, releasing their pollen on the breeze often creating drifting ‘clouds’ of it in the process. (Because of the shared ancestry of plants many characteristics are shared amongst the different groups. It can be confusing. Wind pollination is a more ‘primitive’ pollination strategy, obviously retained by some of the later evolving flowering plants.) No, it’s not very efficient, but it is effective. As a secondary result these wind pollinated plants are very often the ones that we find ourselves allergic to, so bountiful is their pollen upon release. Their prodigious production of pollen also aids scientists in their efforts to ‘date’ various artifacts, objects and fossils they may find that contain persistent pollen grains. (Under examination pollen of Monocots differs by possessing a singe ‘pore’ while that from Dicots has several such pores. This can only be seen under magnification and is of the most interest to those ‘dating’ plant material or sites as the pollen can be preserved for very long periods under the right conditions.)
For botanical purposes a fruit is the tissue that encloses the seed or seeds. It generally ‘ripens’ as the seeds ripen and may then become attractive to animals who would consume and thus carry the seed to other sites where it would be defecated, along with the animal’s waste, and perhaps grow another plant. Some Angiosperms, like the Grasses, pursue another strategy, forgoing the production of pulpy large fruits, instead dropping their seed directly on to the ground below or having their seed eaten and scattered specifically by seed-eating birds and other small animals. Wildlife has evolved along with the plants of their region.
Interestingly Monocots have flowers divided into multiples of three, three petals, sepals, anthers, etc. and their ovaries are divided into three chambers. Several genera may confuse the novice. Lilies appear to have six petals when they are actually petals and modified sepals. Sepals are the outer most vegetative bracts that protect and enclose the flower in bud…only sometimes they appear nearly identical to the petals. In these cases they are often called tepals collectively. If a plant first appears to be a monocot in flower a simple count of the flower parts will usually indicate whether it is or isn’t. Dicots tend to have 4 or 5 petals, though exceptions occur with some forming more congested flowers. These are often selected by nursery people to propagate and sell on to customers for their unique character. Then there are the members of the Orchid Family, another Monocot.
An Assortment of Blooming Monocots
























The Orchids possess ‘irregular’ blossoms. They still have the characteristic 3 petals and 3 sepals, but the sepals have a unique shape and may or may not be green or variously colored. The lowest petal is often modified into a pocket, pouch or spur. They can be bisected into equal ‘halves’, but unlike most other Monocots, they have a top and a bottom portion. In general Monocots have regular flowers with a simple ‘radial’ symmetry. The appearance of Orchids would suggest that they might belong to some other group, but Monocots they are. Their flowers share fewer characteristics with the Dicots. The leaves, however, are very much those of a Monocot. (Orchidaceae are an interesting oddity, made even more so by the fact that they are the single largest plant family in the world including about 800 genera and species, totaling, arguably, between 20,000 to 35,000, depending on the taxonomist, a staggering number, that, altogether, occupy a small fraction of the Earth’s land surface.)
The attempt to classify plants, or any living organism, seems to run into exceptions everywhere. Classifying life is a human endeavor and our knowledge is imperfect. Another family, the Aroids, contain over 100 genera and 3,700 species, which include the Arisaemas or Cobra Lilies, Zantedeschia or Calla Lilies, Anthurium and Philodendron, some taxonomists even include the aquatic Duckweeds. All possess the single seed leaf of all Monocots, but have very unconventional flowers whose structures often defy the 3-part rule and display an inconsistency within the genus, and in some cases, within a single species itself. They possess an inflorescence that utilizes a structure of a modified leaf, a spathe, in a somewhat ‘urn’ or goblet shape, but not always, sometimes ‘hooded’, that can enclose a ‘club’ shaped spadix that carries the actual individual flowers…though many of the more than 3,000 species in the family have no spathe. The tiny individual flowers crowd each other arrayed on the spadix, sometimes with male flowers toward the top, releasing their pollen, before the females, down below, reach their mature/receptive stage, or each flower may be complete with both sexes.
Individual Aroid flowers are stripped down to bare essentials typically having 4 to 9 tiny tepals and 4 to 12 stamens…sometimes more! They are highly variable. The plants themselves have growth habits varying from Philodendron’s, arborescent, tree forms, vines and epiphytes, to herbaceous perennials that grow a ‘modified’ stem, like Colocasia esculenta, a corm, from which Hawaiian’s produce the starchy food, taro, to Wolffia microscopica, an aquatic duckweed, that completely forsakes the spathe and spadix structure, bearing the smallest inflorescence of any plant in the world, just a few millimeters across and Amorphophallus titanium, the Titan Arum or Giant Corpse Flower, restricted to western Sumatra, whose ‘classic’ spathe and spadix form, is the largest ‘unbranched’ inflorescence in the plant world, reaching up to 6’ tall atop its flowering stem! (Here’s a second link, to totally geek out on, to a Cal Berkeley page discussing the idea that plants, as living organisms, should produce a magnetic field, as animals do, through the coherent movement of electrons within them, here tested on the Titan Arum.)
[The honor of the largest ‘branched’ inflorescence goes to the Talipot Palm, Corypha umbraculifera, another Monocot, and, a monocarpic Palm, that can live as long as 60 years, reaching a height of over 80’, before it produces its inflorescence, a structure itself that shoots up another 20’-26’ and contain from one to several million tiny 3-parted flowers! (Clic on this link to visit Louis the Plant Geek’s entertaining and informative posting about this huge and rare Palm.) Monocarpism, the trait of once flowering and then death, is scattered amongst plant families and species. It is generally exhibited by plants with relatively large inflorescences with prodigious seed production, not unlike the annuals many of us are used to…though often of larger scale. Flowering has, after all, its primary purpose in the perpetuation of the species, though there are some angiosperms, flowering plants, that have become almost totally dependent upon the vegetative increase of themselves, spreading from their root structures or their modified stem tissues, that comprise bulbs, rhizomes and tubers, while their flowers produce no or very little viable seed. Horticulturists often seek out oddities like this to bring to market because many consider ready spreading by seed, or their ‘messy’ fruit which may accompany them, in their gardens, to be a ‘negative’.
Leaves: Parallel or ‘Netted’ Venation











Leaf venation can be helpful when trying to identify a plant. Monocots have ‘parallel’ veins. While not strictly and mathematically true, the veins are evenly spaced along the ‘mid-rib’ arcing out towards the margin or edge, often, though not necessarily, curving as they go toward the apex or tip, each maintaining the same distance to the others. There is a uniformity to them. Whether the leaves are narrow like most grasses or more broad and stubby like many Hosta, they possess this same parallel characteristic. The leaves of Dicots exhibit a pattern of netted venation generally with a predominate mid-rib running its length and often with evenly spread secondary veins extending out from it to and ending at the leaf’s edge or margin. It is between these where the ‘net’ forms.
As with all species within a Family or group there will be variation. In the genera comprising Bananas, the veins go straight to the leaf margin, almost perpendicular to the mid-vein, and evenly spaced. In Bananas this may be an adaptation that allows the leaves to shred or tear in windy situations while allowing it to continue functioning photosynthetically, the cells of the leaf still functionally connected to the vascular system. A torn or interrupted vein can no longer contribute to the functioning of the plant.
I have to mention Cornus. Cornus, the Dogwoods, is a genus of Dicots that exhibit prominent parallel venation roughly following the leaf margins while maintaining their individual spacing. In-between these parallel ‘secondary’ veins are a finer, less obvious, network of netted veins. So, on this ‘score’, Cornus spp., would seem to violate the ‘rules’ that divide Monocots from Dicots, but the reality shows how these may ‘grade’ from one to the other. For those of you who want to get your ‘geek on’ check out this page! It clearly illustrates venation and provides links to this dendrology site’s other helpful pages.
Stems and Vascular Systems
One of the most distinct anatomical differences between Monocots and Dicots are their vascular systems and the structure of their stems. Monocot stems have their vascular system, their phloem and xylem, fluid conducting tissues, in scattered bundles within the stem generally with more toward the periphery, the rest of the stem tissue is composed of, non-vascular, ‘ground tissues’ or undifferentiated parenchyma cells and ‘hardened’ fiber cells. Some consider their stems or trunks to be ‘extensions’ of the leaves themselves, commonly packed into a narrow ‘cylinder’ on ‘top’ of which rides the apical meristem. Monocot stems contain no cambium layer. These structures all contribute to a stem’s structural integrity. (Meristem tissue is commonly a single cell ‘thick’ and is composed of rapidly dividing/increasing cells. Unlike animals, that continue replacing their individual cells through out their lives, plants continue actively growing in size until, becoming overly mature, a state determined by the plant’s DNA and its growing conditions, or is compromised by those same growing conditions, disease or infestation, they stop.)
In Dicots the vascular bundles are arranged in a ring forming a ‘cylinder’ just inside the cortex. The middle portion of the stems of annual and herbaceous Dicots, the new growth of extending branches, terminals and of their seedlings, is spongy pith. The outer cortex, is made up of variety of different cells, sandwiched between the epidermis on its outside and an endodermis on the inside. Pore spaces develop in the cortex that serve to store a variety of necessary plant compounds like carbohydrates and starches, resins, essential oils and latex, varying between species. This layer also contains some chloroplasts and is capable of photosynthesis. In Dicots, a cambial layer, the cell forming meristematic tissue, sheaths the entire plant, its roots, main stem and branches. Herbaceous Dicots have cambial tissues that have ‘lost’ the ability to produce secondary growth and so do not add girth, woody tissues, to their structure, their above ground structure dies back and is replaced every year, but they do still contain buds that allow for branching along their stems.
The stems of woody Dicots produce secondary growth annually, from their cambium, which is visible as rings when you examine a cross-section of any part of its supportive structure. This secondary growth forms as phloem and xylem tissue, the xylem, water carrying vascular tissue, becoming the plant’s ‘sapwood’ before ‘dying’ and becoming the static wood bulk, or ‘heartwood’, of a Dicot tree or shrub. A woody Dicot gains strength by adding bulk to the circumference of its trunk. A woody Monocot, such as palms, gain strength by strengthening the tissue it already has.
Dicots Growth and Meristematic Tissue
 Monocots and Dicots both grow through cell division that occurs in their meristematic tissues. No such tissue, no such growth. A plant’s other cells are incapable of doing this. For Dicots this tissue is in a generalized layer inside its cortex and their buds. The meristem grows with the plant, from the tiny apical meristem it began with as a seedling. This pattern is very similar within the stem and root structures. As Dicots grow extending their structure buds ‘ride’ at the tip dividing as they grow and in the process, produce the meristem containing, cambial sheath, that encloses this extension creating additional buds at intervals largely determined by their DNA, along their length that may grow at a later date. If a Dicot produces modified stem or root structures like bulbs, corms, tubers and rhizomes, extensions of the original, these are fully capable of growing into another plant from their meristem tissue as well. Their cambial sheath has somewhat reduced and modified abilities, compared to the apical meristem in their buds and largely are left to produce a particular kind of growth over the life of the entire plant. Monocots don’t do this.
Monocots and Dicots both grow through cell division that occurs in their meristematic tissues. No such tissue, no such growth. A plant’s other cells are incapable of doing this. For Dicots this tissue is in a generalized layer inside its cortex and their buds. The meristem grows with the plant, from the tiny apical meristem it began with as a seedling. This pattern is very similar within the stem and root structures. As Dicots grow extending their structure buds ‘ride’ at the tip dividing as they grow and in the process, produce the meristem containing, cambial sheath, that encloses this extension creating additional buds at intervals largely determined by their DNA, along their length that may grow at a later date. If a Dicot produces modified stem or root structures like bulbs, corms, tubers and rhizomes, extensions of the original, these are fully capable of growing into another plant from their meristem tissue as well. Their cambial sheath has somewhat reduced and modified abilities, compared to the apical meristem in their buds and largely are left to produce a particular kind of growth over the life of the entire plant. Monocots don’t do this.
Woody Dicots grow through the extension of their terminals and by adding to their girth through the ‘secondary growth’ of their cambial sheath. The cambium that encases their stems, branches and roots, divide, duplicating the same tissues that ‘thicken’ them over time. To the inside of this meristematic tissue it produces ‘wood’, the xylem vascular tissues that are comprised largely of lignin strengthened cellulose around the center pith that formed initially, and cortex cells to the outside, which gradually slough off as it ages as old bark. The seasonally changing conditions effect the growth rate of these tissues resulting in the appearance of the characteristic ‘ring’ structure observable in cross-sections of woody Dicots. This same cambium makes possible the formation of branches forming from buds that have previously been lying ‘invisible’ and dormant. The terminal of each branch, its apical bud, is also meristematic, capable of the more complex cell division and the creation of additional vegetative and floral buds.
A similar action happens at the Dicot’s base, from its ‘radical’, which produces a tap root structure that ‘branches’ in much the same way as its top growth, extending, down and out, into the soil, adding girth as it grows outward and producing an ‘endless’ supply of root hairs that grow and are sloughed off. These meristem tissues repeat growth patterns, determined by the conditions and DNA, that result in a characteristic form that an observer can see duplicated, though not necessarily identically, amongst other individuals of the same species.
Monocots Growth and Meristematic Tissue
Monocots are, again, different in their ‘particulars’. Tissues are still grown through cell division, but they don’t produce any secondary growth…and they don’t form secondary buds at internodes along their extending stems, trunks or roots. Everything begins from their single apical meristem, located, initially at the base of the plant, where in some cases it stays atop a more or less static stem, otherwise ‘riding’ atop it as it extends in a way similar to Dicots.

The concentration of apical meristem in Monocots, is responsible for the major structural differences in these two groups of plants. As the stem of a Monocot grows through extension, it produces no cambial sheath. They have no such tissue ‘sheathing’ them, no branches with their own buds, at their terminals or internodes, nor such tissue on their roots or their tips. They do not form annual ‘wood’, even in those plants like Palms, the trunk forming Yucca, and the ‘woody’ stemmed Bromeliads and Irids. These so called ‘woody’ structures are structured in the same manner as the non-woody Monocots, only more densely and much larger, with tightly compressed ‘hard’ fibers and ground tissues, their vascular tissues embedded within them as described above, all originating from their singular apical meristem, in a ‘push’ of growth. Their ‘woody’ stem growth is added annually only at their single stem tip. They extend up with their ‘finished’ diameter. [Read this again and remember it!]
[Please note that there are almost always exceptions to the rule when it comes to living organisms, in this case, the branching of such plants as several Yucca species. This branching must still arise from meristem tissue. A few Palm species may produce multiple stems from their meristem, like the temperate Mediterranean Fan Palm, Chamaerops humilis, but as a rule, they don’t branch above ground, even if you remove their tops. Many Dicots will branch and will do so heavily when a terminal is removed because terminals and branch tips all contain a ‘bud’ containing meristematic tissue. Undamaged, under normal conditions, such buds release a hormone that tends to suppress the growth of dormant or suppressed buds ‘downstream’, hence, when terminals are removed, new branches begin growing below until another bud becomes dominant and begins to release the hormones that again suppress growth below. Monocot stems don’t branch. They have only one terminal that tends to dominate others nearby in a clonal colony springing from the same modified stem structure.]
The above ground growth of a Palm, and other Monocots with woody stems, is very similar to on e another…including members of the Agave family. The apical meristem, contains ‘primordial’ tissues of the plant that divide and grow producing the the stem, its leaves and flowers. These plants use these primordium as a kind of pattern, to grow themselves through cell division, growing all its cells according to its DNA, in response to external keys and conditions including temperature, moisture availability, the amount of energy stored in the form of carbohydrates and day length, growing the stem upward, full size, each leaf forming, expanding from its own primordia, flowering at appropriate times set by the plant itself. These primordial tissues arise from the apical meristem ‘riding’ at the top of the stem as it extends upward. This stem or trunk essentially forms from the base tissues of leaves, connecting each one, continuously, down through the still living tissue of the stem or trunk, to the plant’s roots. They share this characteristic with the grasses and Bamboos, any of the Monocots that form an above ground stem.
e another…including members of the Agave family. The apical meristem, contains ‘primordial’ tissues of the plant that divide and grow producing the the stem, its leaves and flowers. These plants use these primordium as a kind of pattern, to grow themselves through cell division, growing all its cells according to its DNA, in response to external keys and conditions including temperature, moisture availability, the amount of energy stored in the form of carbohydrates and day length, growing the stem upward, full size, each leaf forming, expanding from its own primordia, flowering at appropriate times set by the plant itself. These primordial tissues arise from the apical meristem ‘riding’ at the top of the stem as it extends upward. This stem or trunk essentially forms from the base tissues of leaves, connecting each one, continuously, down through the still living tissue of the stem or trunk, to the plant’s roots. They share this characteristic with the grasses and Bamboos, any of the Monocots that form an above ground stem.

In this cross-section of the growing end of a Palm, LBs are the older Leaf Bases of the Palm, still actively photosynthesizing and receiving water and minerals via the vascular system. DLs are the Developing Leaves. No flower primordia are shown in this photo, though they too would arise here.
Palms, as Monocots, unlike woody Dicots, whose old woody tissues cease growing/changing, die, remain living for the life of the tree, 100 years, and much more in some cases, adding lignin and cellulose the entire time, increasing its strength. The Wax Palms of Colombia’s rain forest, Ceroxylen quindiuense, (here’s a link to an interesting site on the precarious existence of these Palms endemic to one site high in Colombia!) is the tallest Palm and Monocot in the world, reaching heights of 160’- 200’, while the now rare Jubea chilensis, the Chilean Wine Palm, with the thickest stem of any Palm and Monocot, can form trunks 5’ and more in diameter while reaching a height of over 60’ after many years.

This Yucca rostrata was planted as a 1 gal plant 18 years ago the same time the Chinese Windmill Palm behind it was from a 5 gal pot. You can see the emerging inflorescences in the top of the Palm and its old frond bases that remain after cutting. The Yucca has yet to bloom and I’ve left the now adpressed old leaves in place on the stem.
This growth habit gives Monocots their readily observable radial leaf arrangement. These sometimes emerge in a whorled pattern similar to the branching on many Conifers, only these do so only with leaves. Many Monocots, especially those with woody/perennial stems, have leaves that emerge in a spiral pattern, though in the case of many Agave, Yucca and Cordyline the arrangement is very compact because the stem can be relatively short. Some Palms can have their ages approximated by counting the leaf scars left on their trunks, as some produce a kind of banding. The older these are, the taller the stem.
Bulbs, or ‘geophytes’, have a distinct ‘twist’ on this patten. These are modified stems that enclose their apical meristem within carbohydrate storing tissues. Other plants form related ‘stem’ structures in the form of corms and rhizomes. These structures in turn increase extending themselves, rhizomes, or forming more, bulbs and corms, each rhizome extension containing a node with its own apical meristem. Each bulb or corm offset have their own meristem tissues as well. (More on these later. Many Monocots don’t form an above ground stem. There’s remain at or below ground level. Others produce both, above ground stem structures and below ground storage, propagating structures).
Palms and other woody Monocots, generally have an above ground perennial structure, a trunk, and once they reach a mature size the annual cycle of leaves and flowers begins and continues over the plant’s remaining years. Some species of Palm have very truncated trunks that remain underground, e.g., Sabal minor, Rhapidophyllum hysterix and Nannorrhops ritchiana. Other Monocots, like grasses, produce new top growth annually that generally dies back at season’s end and is replaced the following year. Monocots like Agave, that are monocarpic, that flower only once, may quietly grow their leaves at the top of their compact swollen stem structures for as much as 30 or more years before an internal biochemical ‘trigger’ releases an explosive extension of stem growth with the apical meristem ‘riding’ it to the top where it forms a relatively massive inflorescence producing no other basal leaves.
A practical result of this growth pattern is that if a Monocot loses the portion of the top of its stem that includes its apical meristem, if it is cut off, the stem or ‘trunk’ is done. The meristem is removed with it. No bud tissue will be ‘released’ to form a new leader or a ‘bushy’ top from lower down on the stem. No flowers will form. That opportunity was removed with the stem’s top! This is true whether you remove the top of a Lily stem, a Bamboo culm or a Palm Tree. New shoots in many species can arise from another node on a rhizome or bulb, if they are available, but that particular stem is finished…it will play out what remains of its particular life, which might take a couple of years with Palms, without flowering and then die. A Palm’s leaves, are evergreen, but if new growth is halted by such cutting, they can last only so long, before the plant finally succumbs. Having said this….
Intercalary Stem Growth and ‘Exceptional’ Branching

A new Lily Family member in my garden, Polygonatum kingianum, illustrating it stem extension via the growth of its interclaray meristem tissue even spacing its nodes along its lengthening stem. Here flower buds are hanging from some of the nodes and whorls of leaves…without any axillary branching.
Because Monocots are such a broad group, exceptions are every where and no less so in stem elongation. Many Monocots possess intercalary meristem tissues at the nodes along their stems. In forming stems these tissues divide rapidly, elongating the stem tissues between the nodes, creating the internodes between nodes. This allows these plants to gain height rapidly, much faster than if they are limited by depending on a stem’s single apical meristem. Leaves and sometimes flowers may form from intercalary meristem in some species as well. Initially the emerging plant, creates a compacted version of itself, with its nodes tightly together created by the dividing cells of its apical meristem and then, later extending it from the nodes, creating a hollow or solid stem, depending on species. Bamboos, which I discuss separately below, do this, along with the flowering culms of grasses only in the case of Bamboos, this tissue often produces minor branching at the nodes. Agaves, when they switch into flowering mode, do this as well as do most Liliaceous plants, growing solid stems. Some of these may form bracts or attenuated leaves attached to their elongating scales. There are a great many Monocots though that, among them the ‘geophytes’, because they have such shortened stems as mature plants, don’t do this.


Bananas and Their Pseudo-stems








Some Monocots form what botanists call ‘pseudostems’, their stems or ‘trunks’, are the result of tightly packed leaf tissue giving the appearance of a stem. This is not just an issue of semantics. Keep in mind that true stems have a different structure than leaves, that leaves can separate from a stem as they are formed of different tissue. A Banana’s pseudo-stem may be composed of 20 or more tightly held leaves that fan out into thin leaf ‘blades’ at the top of the pseudo-stem where they arch outward, but the veins and midrib of the leaf run continuously, and separately from each other, from the meristem, at the plant’s base, the ‘true’ swollen stem tissue, to the leaf tip. The ‘stem’ of a banana is very short and is limited to the rhizome at or just above ground level. ‘Hardy’ Bananas, in cold enough winters, will freeze down to the bud, their leaves/pseudostems collapsing in a mushy/stringy mass on top. Their bud and rhzome, their modified stem, can remain healthy and firm, waiting for conditions again supportive of new top growth. This is why if you cut the pseudo-stem off above this point, you have only removed leaf tissue and have not cut the ‘stem’. A Banana will continue to produce new leaf growth, growing a new pseudo-stem because the meristem remains undamaged, the primordia, are intact. Many other Moncots rebound from cutting back, because the cut top growth is only leaf tissue. Cutting the leaves from an Iris or the short stemmed grasses we use in our lawn blends does not kill the plant…it only delays the eventual flowering. The Banana is not an exception among the Monocots, it does, however, require that we examine it more closely before we act.
Bananas, Agave and other Monocots may have the ability, when under drought stress, to ‘pull’ moisture from their leaves to help sustain the more vital tissues within their rhizome and meristems. These softer, more temporary, above ground structures are sacrificed for the health of the overall plant. This enables these plants to survive drought conditions their leaves shriveling in the process. This is a characteristic that gardeners take advantage of when wintering over less cold hardy Monocots, bringing them in to the cool dark, protecting them from cold extremes, while allowing the plant to grow another day. With this treatment Bananas in fact shrivel like raisins in cool dry storage protecting their apical meristem. If the temps are too high when stored this way the Bananas metabolism remains higher and the plant continues pushing new growth, transpiring a lot of water that it is drawing from itself and by doing so weakening it by spending its carbohydrate reserves as well as through loss of water
The modified stems and meristem of Monocots, before they can produce mature top growth, must grow ‘itself’, sometimes utilizing juvenile leaves, until it is capable of growing a mature structure above ground. The Colombian Wax Palm mentioned above, spends it’s first 15 years growing its base, meristem and roots, before it begins to add significant vertical growth to its trunk. A mature meristem produces mature growth capable of flowering. Think of a little Lily bulb scale that must grow into a mature bulb capable of flowering…this will take several years. When begun from seed, Lilies take 9 years +or- before their bulb is big enough to produce a mature flowering stem….In its immature years it produces growth shorter and smaller, that in its youngest stages, may not even resemble that of the mature plant, stems incapable of flowering…while they are, at the same time, producing more bud scales, or other stem structures, that they nurse along, again forming from the basal meristematic tissue. Once fully formed, it doesn’t grow anymore broad.
Palm trees continue growing each year adding height, each year producing another cycle of flowering. The top most portion of a Palm’s growth is in a ‘spear’ form as it emerges. Each new frond’s petiole ‘moves’ to its final position along the sides of a trunk’s cylinder as it extends its vascular and non-vascular tissues. These form a consistent pattern of dead petioles or leaf scars along the length of the trunk. After a few years an entire frond is abandoned, dries and loses its ability to both aid the plant photosynthetically and to receive the water and nutrients it needs to continue in an active state…some species will later shed them. The meristem has literally moved on above them. Over the mean time, additional spears/fronds form steadily adding to the plant’s overall height.

A Palm with a distinct crownshaft, showing the prominent leaf scars where they fall/tear away. The inflorescence emerges vertically at this point before branching and displaying its little flowers. Taken at a friend’s in Shell Beach, CA…not Portland.
Some Palms form what is called a crownshaft. (This link connects you to a very helpful forum discussing Palms…on this particular page a member discusses just what a ‘crownshaft’ is.) This is an elongated structure at the base of a leaf where it attaches to the trunk. These partially ‘wrap’ around the trunk forming a smooth contrasting surface near the top of a trunk. When one of these leaves are shed the entire base peels away resulting in a distinctive leaf scar at the point from which it initially emerged from the trunk, now below the rising still growing trunk. Their inflorescences emerge at this point along the trunk appearing as little ‘arms’ from the apical meristem formed from beside the leaf forming center portion of the apical meristem. This all important tissue is not at the trunk’s apex, but below it sending growth up and out. This location also provides it with cold protection shielded within the other surrounding tissues.
Meristematic Tissue and Flowering
Monocot flowers initiate along with the stems from their apical meristem. In Dicots, meristematic tissue is located in the cortex/cambial tissue that sheaths the plant. While buds, vegetative and floral, form at locations around the plant, typically at branch terminals and leaf axils or, in the case of plants like Cyclamen and Asarum, the flowers emerge from the top of their flattened modified stem tissue. (And, yes, their are a few trees, woody Dicots, that form flowers directly on their trunks, mimicking their Monocot geophyte cousins.) All growth is expressed through cell division, the addition of one cell to the next, each of the correct type, produced as needed, precisely where it needs to be. Flowers are special organs composed of relatively complex tissues. They are not the ‘simple’ extension of stem tissue, or of leaves that may be added as stems elongate. Flowers are more rare structures that appear in limited and particular locations, often at a particular ‘moment’ when conditions are met. A Monocot’s apical meristem, as pointed out above is associated with its stem tissue whether it is modified into a ‘storage organ’ as a bulb, corm or rhizome or forms an aboveground stem or trunk. Flowers form, therefore as direct extensions of these tissues. Whether Palms, Bananas, Grasses, Orchids, Tulips or any other other included family.
 Think of the common Tulip. Tulips produce bulbs and belong to a larger group of plants known as geophytes. Geophytes are plants that survive part of their annual life cycle as a dormant, fleshy underground structure, the mediterranean geophytes, ‘resting’ during the dry summers. Types of geophytes include bulbs, corms, tubers, tuberous stems, tuberous roots, rhizomes and pseudobulbs…but Tulips don’t sit idly by over Winter. While seeming to be dormant, they are busy producing a tiny primordial ‘plant’ within the bulb from its apical meristem, connected to its basal stem, that is actively consuming energy stores from the bulb. When Spring arrives, soil temperatures warm and the already ‘formed’ flowering stem, the scape, erupts from the bulb, powered by the stored starch enabling the rapid growth of flower and leaves before the plant can begin photosynthesis. From the base of the bulb, a broad, initiation zone, the underside of the meristem, roots extend out and downward. The scape serves to support the flower and transports the sugars, proteins and other components needed to grow it, the flower and its later seeds. Each species of bulb follows its own pattern with many of our temperate Spring flowering bulbs replacing their spent carbohydrates and the growth of new bulbs in the cool Spring weather after flowering. For many of these a ‘hot’ spring shortens this period and signals the plant to begin the cycle for the next year, which will have smaller bulbs, as a result of a shorter photosynthetic period, that flower less vigorously. Presumably, if a bulb’s growing conditions do not trigger the proper signals it may ‘skip’ a year. Each species will have its own ‘preferred’ conditions to produce optimum growth. Bulbs like the Tulip are the ‘simplest’ case in terms of flowering for monocots though even they vary somewhat in how they do it.
Think of the common Tulip. Tulips produce bulbs and belong to a larger group of plants known as geophytes. Geophytes are plants that survive part of their annual life cycle as a dormant, fleshy underground structure, the mediterranean geophytes, ‘resting’ during the dry summers. Types of geophytes include bulbs, corms, tubers, tuberous stems, tuberous roots, rhizomes and pseudobulbs…but Tulips don’t sit idly by over Winter. While seeming to be dormant, they are busy producing a tiny primordial ‘plant’ within the bulb from its apical meristem, connected to its basal stem, that is actively consuming energy stores from the bulb. When Spring arrives, soil temperatures warm and the already ‘formed’ flowering stem, the scape, erupts from the bulb, powered by the stored starch enabling the rapid growth of flower and leaves before the plant can begin photosynthesis. From the base of the bulb, a broad, initiation zone, the underside of the meristem, roots extend out and downward. The scape serves to support the flower and transports the sugars, proteins and other components needed to grow it, the flower and its later seeds. Each species of bulb follows its own pattern with many of our temperate Spring flowering bulbs replacing their spent carbohydrates and the growth of new bulbs in the cool Spring weather after flowering. For many of these a ‘hot’ spring shortens this period and signals the plant to begin the cycle for the next year, which will have smaller bulbs, as a result of a shorter photosynthetic period, that flower less vigorously. Presumably, if a bulb’s growing conditions do not trigger the proper signals it may ‘skip’ a year. Each species will have its own ‘preferred’ conditions to produce optimum growth. Bulbs like the Tulip are the ‘simplest’ case in terms of flowering for monocots though even they vary somewhat in how they do it.
Other bulbs, follow a similar cycle, flowering in Fall and are followed by vegetative growth to renew their bulb, a strategy that will obviously only work in milder climates. Others bloom in the Summer after the foliage grows up. The flowers of some, like the Summer blooming Crinum amongst the members of the Amaryllidaceae, lead with their foliage first, their scapes curving upward from the base of the bulb rather than emerging from the top and their thicket of foliage.
A ‘true’ bulb, like Tulips and Lilies, are composed of a single very short section of flattened stem containing one node and producing one flowering stem. The bulb ‘scales’ enclosing the center are modified thickened leaves packed with carbohydrates. Over time these can increase by offsetting, the ‘scales’ growing from axillary buds, including their own bit of meristem grown from the mother plant. Cut or break off this singular flowering stem and there will not be another one. Remove its foliage too soon and the bulb cannot replenish itself.
Corms may often look like ‘true’ bulbs, but they are not. Some, like those of Eucomis, begin from a longer section of stem with one or more nodes and take on a more irregular shape as they age, growing and adding nodes to their bulk that will form multiple crowded flowering stems and sets of leaves. They are solid starchy structures when cut into not the layered structures of true bulbs. They don’t produce scales from which they can be increased. Instead, they can be cut into pieces that will grow on successfully if they include a bud or ‘eye’. While other corms will actually form a ‘stack’ of young corms on top of the older ones…think Crocosmia.
There are other modified stem structures that together comprise the geophytes, adding fibrous stems, tubers and rhizomes to bulbs and corms, all following their particular pathways to flowering, with many similarities. The Iris genus varies between species some forming true bulbs like Tulips others growing and increasing from rhizomes. Some, like Iris unguicularis, issuing from a rhizome, bloom in Winter with their scapes emerging above their tough leaves, and do so successfully here in Portland, where temperatures may drop below freezing for periods, but not too severely to disrupt the process. Many bulbs have alpine origins and ‘require’ a substantial cold period. All of these Monocots, commonly and collectively known as ‘bulbs’ begin from their apical meristematic tissue at their bases.

An Orchid with a sympodial pseudo-bulb. Wikipedia
And then of course, just to screw things up even more, many epiphytic and terrestrial Orchids form what are called, ‘pseudo-bulbs’, above ground structures composed of a single, stem internode thickened with starches, meristem at their base, flowers and leaves arising directly from these…placing them outside the ‘realm’ of the below ground geophytes. These pseudo-bulbs can form into chain-like spreading structures, termed ‘sympodial’, the oldest declining upon completion of flowering and seed formation. Another variation of this, ‘stack’ these, each internode/pseudo-bulb, one atop the other, ladder like, each with its own leaves and flower. What’s important for the gardener to remember is that these aren’t true bulbs and that there is no appropriate depth to set them out in the soil. This will cause them to rot. While true bulbs have their roots form similarly at bulb’s base, these won’t tolerate such treatment. Similar doesn’t mean that they’re identical.

Calypso bulbosa. I found only two tiny groups of these growing on the north facing slope of the Columbia River Gorge on the trail below the Memaloose Overlook…no where else in the Park.
Calypso bulbosa, a rare native Orchid, almost anywhere it can be found growing across its broad, northern temperate Arctic range, is one of the pseudo-bulb forming species, though it is often described as a corm….Alas, others don’t distinguish between these two structures. Such confusion can lead gardeners into difficulties with growing Orchids successfully, which is not necessarily the case with Calypso bulbosa. This Fairy Slipper Orchid will defy your efforts to grow it in the garden. Internet sources often list this species as a denizen of northern boggy, wet, habitats, rather incompletely, because its ‘preferences’ are more particular, often occurring on dryer humus rich hummocks above the surrounding swamp. Locally, I’ve found these in the Columbia River Gorge growing in thick humus layers above quick draining, shattered basalt, with a northern or shaded aspect, or simply beneath a forest canopy of mixed Douglas Fir/Big Leaf Maple. They seem relatively well adapted to our dry summer climate. Areas where they are found in the PNW have consistent rainfall throughout the Fall, Winter, Spring season, after which, these sites can be quite dry. They seem to have a requirement for ‘cool’ soils and grow in very particular ‘soil associations’ on their sites. This above ground adaptation may be a response to cool, wet, conditions of their region. A great many Orchid species are native to tropical and sub-tropical areas, it would be interesting to see if there’s any correlation between this and their ‘bulb’/root structures.
All Monocots that don’t form true stems or trunks, flower in some variation of the above. Regardless of it length or modified structure, the meristem sits atop it. Members of the Bromeliadaceae, which may be either stemless or stemmed, send their inflorescences up from their meristem following a route between the ‘hard’ clasping leaves, emerging from the center ‘bowl’ in many like the genera Neoreglia and Aechmea.
Plants like Iris, herbaceous perennials, that grow from a perennial and extending rhizome, form new above ground tissues, leaves and flowers, from a ‘bud’ or ‘eye’ on this section of rhizome abandoning the old. Older rhizome sections, though still alive, no longer have active meristem tissues. For many of these plants, successful flowering is a trigger, that precedes the ‘death’ of the supporting meristem. The rhizome, ever extending, creates new flowering possibilities over time. A similar thing happen with many bulbs, with each bulb maturing, flowering, and then succumbing to be replaced by the offsets, cormels or bulbils, produced by the ‘mother’ bulb. The mat forming Bananas of genus Musa do a very similar thing, each stem that successfully flowers, signaling the bud on the rhizome, to shut down when its ‘finished’. While propagating these rhizomatous Monocots, the old rhizome should be cut away as it won’t produce new growth under normal conditions.
People who are unfamiliar with Palms are often startled to see where the inflorescences emerge on the trunk, below the terminal and sometimes from several feet below, at the base of the crownshafts which develop on some species. Like any other flower they are initiated and grow from the meristematic tissue in a Palm’s apical bud, which in the case of Palms, perhaps, can be located several feet below what appears to be the tapered top of the trunk. From it issues out the spears vertically that expand into the new leaves and, just off from the center arise the inflorescences, vascular bundles and trunk tissues forming in this zone as well separating the leaves and inflorescences in a pattern characteristic to the species.

The bud atop the flowering stem of my Manfreda undulata ‘Chocolate Chip’, with the leafy bracts forming from the meristem, elongating and then being left behind along the lengthening stem in a precise pattern…flower and leaf primodia forming within! This skinny stem will reach 4′! (The aphids and their cast off ‘skins’ cling to the underside of the bud, avoiding the rain.
There are more variations on this pattern amongst Monocots. As mentioned above are the genus Agave members which live for several to many years before flowering once, expending their stored starches producing a tall inflorescence, many to well over 20′ tall…their apical meristem, ‘abandoning’ its old position and task of adding to its basal rosette of leaves, ‘riding’ the flowering stem skyward. Other genera in the Agaveaceae aren’t monocarpic increasing the mother plant and flowering year to year upon reaching a mature size including the Dasylirion, Furcrea, Yucca, Nolinia, Hesperaloe and Manfreda among others. All of these share a common structure of their inflorescences, though they tend to be smaller than their Agave cousins who are ‘one and done’.
Bamboos are another case with new culms forming annually, carried in an active living state for several years later dying in a cycle of growth until the plant begins its flowering, often many years later. Uniquely, it is often thought that all of the clones of a particular selection, worldwide, flower simultaneously, producing large amounts of seed, and the plants themselves all dying. Grasses, if the conditions support it, flower annually on structures replace every year.
Roots and Storage Organs
Dicots have a radical, the original seedling ‘root’ that contains meristem. Monocots have ‘aborted’ theirs leaving themselves instead with a general, ‘expandable’ initiation zone where roots form individually, largely unbranched. Monocot’s roots form adventitiously. Each root grows from it and, when it dies and is sloughed off, a new one forming to take its place…adventitious because there are no visible buds from which the growth appears to arise. In many Monocots, like Bananas and many bulbs, corms and rhizomatous plants, the roots give the appearance of a rag mop. This results in a relatively compact soil volume penetrated by a given plant’s roots, though some Palms can spread their roots 50’ out. Each species varies. Bananas are a good example. This can be a great advantage when attempting to transplant or move such a plant in for winter protection. The gardener need not be as careful of the roots of Bananas among others. Typically when replanting a winter protected Banana back out in Spring, you just dig a hole of adequate depth, keeping the base of the pseudo-stem just above the soil line, its starchy rhizome just below…new roots will arise from the base as soil temps warm enough to encourage growth, anchoring and providing the plant with the water and nutrients that it needs.

Monocots can require a lot of time and energy to grow their rhizomes and meristem tissues. Young Palms are slow to establish. They spend several years growing their bases and their meristematic tissue, broadening with it, while producing more roots that serve to anchor and buttress it firmly to the ground before it puts significant energy into lengthening its trunk. In the photo left, the pen, locates the point at which the plant began its growth as a seedling. In the case of Palms this to is the slowest stage of growth. During this period the new plant grows very slowly upward while adding girth, producing the characteristic ‘V’ form shown in this cross-section. They top-growth appears ‘stubby’ in youth before they finally begin to extend and, remember, because they have no cambium tissue, will add no additional girth to their trunk over time. Keep in mind that optimal growth during these establishment years is important because you are growing the base of your Palm. (Sometimes you’ll see Palms with trunks that seem to randomly form narrow ‘waists’, this is the result of stress that reduced the plants vigor over that period. These ‘waists’ will remain for the life of the Palm.) Secondly, large old Palms appear to ‘buttress’ themselves by producing a roots above the soil line that angle out effectively bracing the tree.
All plants have tissues that can store energy in the form of carbohydrates for later use or to ‘power’ the quick growth of asexually produced ‘offspring’. Monocots often do this in the form of storage organs that are the result of modified stem or root tissue. Bulbs, corms, stolons and rhizomes are some of the more common structures used this way by Monocots, though it is also a strategy used by Dicots. (Roughly 10% of Monocot species form woody above ground structures which can store this. The percentage is much higher among Dicots, around 50%.) Carbohydrates tend to be stored in perennial structures. Amaryllis, Lilies, Tulips, Iris, Glads, Eucomis…there are many, many native and ornamental bulbs that belong to the Monocots. These storage organs give these a jump start, pushing a plant from zero to flowering in very short order, followed by a recovery period as the generally later emerging foliage rebuilds the energy stores of the organ.
Many of the these geophytic Monocots pursue the strategy of once flowering and then death…called monocarpic, one time seed producers. After each bulb is spent, ‘daughter’ bulbs are already for following years. They form their offsets, scales, bulbils, in addition to seed, which will ‘nurse’ alongside the large ‘mother’ bulb before they are large enough to flower. Some, like the genus Agave, may take 10, 20 and 30 years or more to grow their stores large enough to power their flowering process and die, producing several hundred flowers when they do. Staying with the Agave, there are those that are very shy to form offsets and others still that don’t at all relying entirely on seed to perpetuate themselves. There are several Agave species, and these maybe the result of thousands of years of selection and cultivation, that form viable plants high up in the structure of their inflorescence that latter root and grow when the structure topples.
Other Monocots form rhizomes, that can spread very slowly forming a tight clumping structure while others can grow quickly into surrounding soil forming extensive colonies of identical clones. Rhizomes, that spread just below the soil’s surface, as well as stolons, that grow similarly above the surface, are extensions, that form nodes that contain meristematic tissue from which roots and new shoots emerge as in Bamboo, Grasses and rhizome forming Irids (Some Irids form bulbs.) These rhizomes are extensions linked directly to the mother plant’s meristem. Each successive extension begins with the preceding section’s meristem. Over time, the nodes on older sections of rhizome lose vitality and quit producing new growth. When making divisions or transplanting these, older sections should be discarded. While cultivating in the garden these are commonly cut, and if vital sections are left behind, can grow into an identical plant, from those pieces that include nodes. This can make the complete removal of these plants difficult. The same happens with bulb forming plants when work leaves the tiny bulbs, bulbils and seed behind. If pieces however are left that don’t include a node or any meristematic tissue, not new plants can form. This is why very often as we work to remove certain plants, we may think that we are successful, but having left viable bits and propagules, they seem to reappear magically and frustratingly. This is true for Grasses and Bamboos as well. Segments without adequate meristem tissues, will not grow.
Dicots, however, because their meristem tissue sheaths all of the roots, in some species can produce adventitious roots and shoots almost anywhere. My experience with the little European Wood Anemone, a Dicot, is that even little bits of broken or cut rhizome, can form into a new plant. Many Dicots can have root, stem or even leaf tissues, induced to produce root and shoot growth by propagators, not just because these tissues cells contain all of that plant’s DNA, all plants do, it is related to the growth generating meristem tissues in these plant parts. Again not all Monocot tissues are capable of this, at least without some effort to induce such growth.
Grasses, Bamboos




I have to say something about the Grasses and Bamboos, a large group among the Monocots and one whose populations cover a large portion of the Earth’s surface. Both of these groups of plants occur as ‘bunchers’ while many more are ‘runners’ and others still are somewhere in-between and in the case of bamboos like Chusquea species are known as ‘loose clumpers’. I’ve already mentioned the apetalous flowers of these, but I want to discuss some of the differences between these two large groups. Their structures are very similar. Each grow with their shoots or culms arising from buds located at the nodes, or joints, along their rhizomes. Their roots originate adventitiously, or spontaneously from an ‘initiation zone’ on the node in a manner similar to all Monocots. No roots or shoots form from the internodes that separate these growth points, though clumps can grow in a dense congested form because the internodes are so short.

This is a section of rhizome from my Phyllostachys aureosulcata ‘Spectabilis’ that had escaped across a sand trench and was extending under my stone patio. A culm is forming from a node near the finger-tip of the glove, growing horizontal. It was just inside the edge of the patio. The bare rootless nodes were under the stone. The heavier rooting was in sand.
The runners among these, whether a timber type running Bamboo or a weedy ‘running’ grass like Quackgrass, send up their vertical growth like any other Monocot. Each shoot or culm, is almost an entity in itself fed or supplied by the entire rhizome/root system and capable of being separated and growing independently. In the case of many Bamboos one must be aware when making divisions that enough rhizome is include to sustain a separated plant. Many running grasses on the other hand can be very good survivors and can grow from a small piece of rhizome containing only a single node.
Once a shoot or culm initiates, if it is broken or cutoff it is not just interrupted, it will see no more growth or extension. Cutting off a grass or bamboo’s culm leaves a ‘raw’ end, because its apical meristem has been removed. You see this far too often in poorly maintained commercial and institutional landscapes. Most grasses and their relatives will only ‘recover’ when new culms or shoots initiate and hide the cut ends. Bamboos are somewhat different.
Grasses and Bamboos, as characteristic Monocots, don’t branch along their length for reasons discussed above. Each shoot, in a sense, has only so much ‘length’ in it. Each shoot extends to its ultimate length determined by its DNA and the vigor of the bud from which it emerges, relatively quickly in spring. Flowering stems may extend later in the season rising above their foliage. These stems and culms do not lengthen over the entire growing season or from year to year. Most of the shoots on a given plant will reach a uniform length or height. Others, smaller in diameter and with less vigor, will be considerably shorter and weaker If cutoff, a shoot or culm does not ‘recalibrate’ and finish by growing out to its intended length, nor do adventitious buds initiate from the cut end or nearest nodes creating a stalkier plant that will flower at a lower height, like many Dicots will. (Some ornamental Dicots are commonly cut back to shorten the lengths of their flowering stems, preventing them from flopping. This doesn’t work with grasses, bamboos and the other Monocots.) It isn’t ever going to flower if cut off. It’s done. Any flowering will come from a new shoot that arises from the rhizome or crown.
Many of us think of our lawns when we thing of grasses. Here in the maritime Pacific Northwest the most common component of such lawns is Perennial Rygrass, a cool season grass that has been bred and selected for its short stature, appropriateness for our soil and climate and its ability to respond to repeated mowing. A good lawn grass has a short, fine, stem. When mown other buds are ‘released’ below to grow thickening the turf.
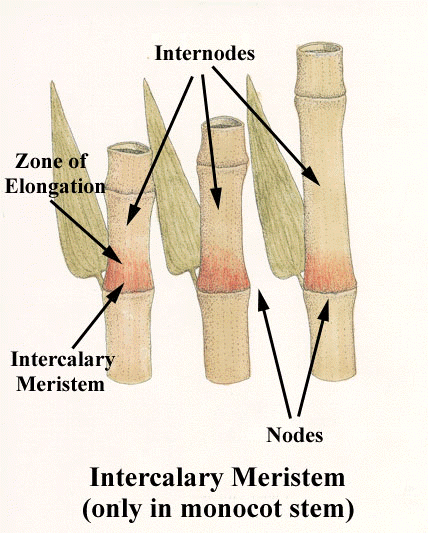 Contrarily, many Bamboos do ‘branch’ at their nodes, each species having a characteristic number and pattern to them. In a Bamboo the nodes contain intercalary meristem tissues, like other Monocots, making this possible (Meristem tissue is identified by where in the plant it occurs). Some like the aforementioned Chusquea can get quite ‘bushy’ adding additional ‘branches’ in subsequent years. All members of Phyllostachys form two branches per node one generally larger than the other. This meristem tissue is similar to that in its below ground rhizome which forms roots and initiates culm growth. The nodes of grasses this meristem tissue, but don’t generally have the capacity to branch. (Exceptions again. Arundo donax, Giant Reed Grass, has a more perennial above ground structure, and if it survives our Willamette Valley winters, will tend to branch somewhat from its nodes, creating a much more congested plant. I choose to cut these down to force a less congested, more elegant structure.)
Contrarily, many Bamboos do ‘branch’ at their nodes, each species having a characteristic number and pattern to them. In a Bamboo the nodes contain intercalary meristem tissues, like other Monocots, making this possible (Meristem tissue is identified by where in the plant it occurs). Some like the aforementioned Chusquea can get quite ‘bushy’ adding additional ‘branches’ in subsequent years. All members of Phyllostachys form two branches per node one generally larger than the other. This meristem tissue is similar to that in its below ground rhizome which forms roots and initiates culm growth. The nodes of grasses this meristem tissue, but don’t generally have the capacity to branch. (Exceptions again. Arundo donax, Giant Reed Grass, has a more perennial above ground structure, and if it survives our Willamette Valley winters, will tend to branch somewhat from its nodes, creating a much more congested plant. I choose to cut these down to force a less congested, more elegant structure.)
Bamboo culms initiate from an apical meristem on a rhizome’s nodes. Anyone who has grown or observed Bamboos closely understand how quickly they can grow from ground to full height, growing several inches and more a day in Spring. The primary purpose for the intercalary meristem is culm elongation. The culms form from their apical meristems in a complete, but compacted form. Upon emerging the culms begin to elongate in a coordinated manner, ‘telescoping’ from each of its many nodes. The intercalary meristem begins rapidly dividing creating the internodes that separate each of these. All of the cells are formed in the process and are quite ‘crisp’ and fragile, becoming ‘reinforced’ later. Some botanists attribute this strategy to the much older family of Horsetails.
Bamboos have a perennial above ground structure, that can live several years, unlike most true grasses that each year abandon the previous years growth giving way to the new. Consistent with this, Grasses flower annually, Bamboos do not. In fact Bamboos can take many years before they flower and then the entire plant dies. It is said that all clones of the same Bamboo, no matter when they are started, will all flower and die at the same time, prompting nurseries to grow on the resulting seedlings and select out the favored cultivars to begin the next cycle.
I should also add that Grasses and Bamboos have a different stem structure than many of the other Monocots, in that they their stems are hollow instead of being filled with parenchyma and sclerenchyma (hard) cells. Bamboos apparently concentrate their fibers and the much ‘harder’ sclerenchyma cells in a thickened ring that, after the initial extension is hardened with lignin and silica making it very durable. (Bamboo shoots, the newly emerging culm tips are often consumed in Asian cuisine as they are ‘crisp’, not hard.) Emerging shoots are subject to breaking so work and casual activity should be limited during this period. In general most grasses do this minimally as their above ground structures, their culms, are replaced every year. A handful of Bamboos, commonly used in construction, are nearly solid and extremely durable like the tropical Guada and a handful of asian species which are the structural ‘timbers’ in my garden gate and framing members of my garden pavillion. The world of plants, however we attempt to classify and structure them, are full of species that don’t fit precisely in one group or another…a situation that should not be a surprise to us as plants have a time tested shared ancestry that was not created to be easily understood by us. There will always be the ‘rule’ and the exceptions.
Conclusions
The point of all of this is that Monocots, by sharing these particular characteristics, become identifiable as such and, by understanding these, they can help improve our practices in the landscape. If we forget this in our cultivation of them, we do so at the plants peril. Also by understanding that a plant is a Monocot or a Dicot we are able to place them into a larger group that makes it somewhat easier to remember because we don’t have to know each plant simply as an individual. An angiosperm, a flowering plant, fits into either group. When we hear the term Monocot or Dicot and think of a plant with one or two seed leaves, it may seem to be a bit of an arbitrary distinction and one that disappears shortly after a plant’s first ‘true’ leaves form, but this is not the only distinction, as I hope I have pointed out above. Botanists had to call them something, so they separated them based on the most basic shared characteristic. Now, through DNA, these links and connections are being either disproved or strengthened, effecting how we ‘draw’ the phylogenetic tree.
For me the over riding difference between these two ‘clades’ is the location of the meristematic tissue and how that determines the way a plant grows. There is a degree of ‘wiggle’ room in the others, some blurring that occurs between the two, for example the above mentioned Asarum and Cyclamen genera, until we remember that ‘geophytes’ occur both Mono and Dicots. The seed leaves are a constant and observable difference between these two when examining seeds and germination, as is the presence or absence of cambium, also known as lateral meristem. The rest are strongly ‘suggestive’, but require that we look at them as a whole. Botanists use very specific language when they talk of their subjects. It may be off putting to many, but it is the only way to really discuss the details of an organism. We have to use language whose words share very specific definitions to give us the precision needed. Such a ‘language’ also opens up to us the amazing diversity and nuanced responses that plants have developed to their local environment and how they have adapted over millions of years. Precise language permits science. Without it we are left with opinion and are unable to examine or evaluate our own, or someone else’s, observations or claims. It gives us a window through which we can watch and learn.
Sources
“Botany in a Day: The Patterns Method of Plant Identification”, Thomas J. Elpel, HOPS Press, 2013. I strongly recommend this book to gardeners. It is a great resource to help you learn the basic characteristics of Plant Families and to better understand what you are growing and see in the landscape. Not to be read in one sitting, but as reference and to help you ‘sort’ out one plant from another.
While working on this posting I came across this page and feel that for those of you particularly interested in Palms, it is an excellent introduction. When putting this section together I realized that I’ve never acquired a book on Palms. I’ll have to correct that. I do a lot of research on the internet. A little suggestion here, whenever I search for specific information on plants, I include in my query the word ‘botany’ as it tends to exclude a lot of sites that just pass on information without checking it first. Footnoted research is indicative of someone doing ‘science’.
A friend gifted me with a copy of the book, “The Complete Book of Bananas”, a self published title by W.O. Lessard, 1992…for an admittedly small audience, especially in the maritime Pacific Northwest, but….
I’ve read several books on grasses and refer most often to: “The Colour Encyclopedia of Ornamental Grasses: Sedges, Rushes, Restios, Cat-tails and Selected Bamboos”, Rick Darke, Weidenfeld & Nicolson, London, 1999.
If you only have one reference on a group of plants, it should be a good one, of course which one is best is arguable. I have Ted Meredith’s, “Bamboo for Gardens”, Timber Press, 2001, and find it more than adequate for my needs. In the case of Bamboo, Portland is also home to one of the more active branches of the American Bamboo Society and there are scattered public plantings and a collection being actively cared for at the Hoyt Arboretum.
I own “Agaves, Yuccas and Related Plants; A Gardener’s Guide”, Mary & Gary Irish, Timber Press, Portland, 2000, and find it very informative. A more complete reference is, “Agaves of Continental North America”, Howard Scott Gentry, University of Arizona Press, 2004.
“Aroids: Plants of the Arum Family”, 2nd ed., Deni Brown, Timber Press, Portland, 2000, I pull out periodically to reference, covers the many Cobra Lilies, Ariseama, Callas Lilies and the many other genera and species of the family that have flowers, looking like they must belong to a plant group other than the 3 parted Monocots.
Good bulb books abound. I have a couple of smaller titles covering the more common bulbs and more recently picked up, “The Color Encyclopedia of Cape Bulbs” Manning, Goldblatt and Snijman, Timber Press, 2002, a book that examines the most diverse bulb region in the world, in South Africa…often to just drool over and wonder about.
There is are many titles covering the various Monocotyledonous Families and Genera, from very technical monographs, to books suited more for the general gardening public. Pick a Family or larger group of plants of particular interest to you and check out a book on them at your library, or if you’re fortunate enough to have a bookstore that carries a wide selection, and peruse them. Don’t just buy the first one you see. It will do you no good if it just sits on your shelf. Most of these aren’t intended to be read in a single sitting, but to be returned to, time and again, to answer questions that will inevitably arise. I find myself to be a bit of an incremental learner, building up my knowledge base along with growing experience. A ‘good’ book is one that will draw ‘you’ back to it time and again for reference unless of course you’re one of those rare individuals with an eidetic memory. Your efforts may be challenging, but they should be rewarding and further your enjoyment with this gardening thing that we do.
Gymnosperms…A Whole Other Group of Plants & How These Fit Together
This is a link to a site discussing seeds, extant and extinct, in some detail, relatively ‘approachable’ of the major groups, with great illustrations.