






Understanding the New Phylogeny of Angiosperms, part 2
We tend to think of evolution as a historical process, something that occurred in the past which has resulted in life today, with us at the pinnacle. Humans with our opposable thumbs, our relatively high ratio of brain to body mass, our consciousness…our souls, we often argue, are the ultimate life form. We have a hard time imagining that this is not the case, that we as a species, are a part of a continuing process, that some day will fade from the Earth, as other species, more evolved and complex, develop. This is what happens to organisms over seemingly impossible long periods of time. It has happened and is still happening to plants. It won’t happen today or tomorrow and this doesn’t mean that what we are or what we do doesn’t matter…because in evolution…’everything’ matters.
Imagine a still-life, a floral arrangement in a vase on your table, carefully arranged, perfect, static….It’s in a state of decline. Each stem has been removed from a living, growing, plant in your garden or by an industry that supplies your florist. When most of us look at the living green world outside we see a ‘snapshot’ of what is there, a single moment in time, incomplete and selective, not unlike the arrangement in a vase. There is something about our vision, our attachments to moments, to these abstract ‘slices’ out of time that we recall, that pulls us out of its continuous flow. Life occupies every moment flowing from one to the next seamlessly, but we tend to remember in fragments. Only if we spend enough time watching life, playing music, dancing, creating works of beauty, do we learn more about the flow of life and time.
When we were young children we were capable of awe, of being struck by the wonder and magic of the life that surrounded us, drawn into it. For many of us as adults, it is these vignettes, the snapshots of memory, that we are drawn to and recall. These can become what we look for, what we see…more ‘real’ than the actual dynamo of life occurring around us. For many, if not most of us, the awesomeness of nature gets driven away by school lessons and the never ceasing assault of messages and expectations of the society within which we live that diminish and demean the world of nature. Some of us, however, remain engaged, enraptured even, by it…open to it. For us we harbor within us that kernel of interest that pushes us to understand this strange and incredibly complex and wonder-filled world. Others of us are struck, at some point in our lives, as if by epiphany, and brought back to this state after spending a number of years trudging dutifully down our chosen paths.
The world, we find, has never been ‘fixed’ in time. The moment that once gripped and defined our attention is blasted apart and we realize that there has been an infinite number of such ‘moments’ stretching back through time, each one connected to all of those preceding it and pointing toward moments still undefined in the future. We discover that life isn’t a string of isolated moments at all, but is rather a constant unfolding of what is and what is to come. Some say that life is the state of becoming! that it is forever unfulfilled, that every moment gives us the opportunity to look about to see again that which we are a part of. In our attempts to comprehend it, to understand it, we necessarily simplify it, make generalizations, so that we can ‘get on with’ our own lives. To do this too much, however, is to devalue and demean it, to limit our understanding and connection to it…our place in it. To understand where we are or where we are going we must understand where we have been as well as the forces that have been moving us along our path.
Botany is part of our story as the human species. Plants are an undeniable link, multiple chapters in our own story. Their origination and evolution is part of us as well. In the world of botany, phylogeny has become the ‘story’ of plants, and to understand it and what systematic botanist are doing today, we have to look back into the intimate connections between plants, of life and its evolution here on the Earth.
Evolution is a vast and controversial topic on which many books have been published. I’ve struggled with some of them becoming overwhelmed with their technical jargon and their scope that covers billions of years. My intent here is to tantalize you, to get you to begin to look at what can be a daunting subject, a subject that is often said to be in conflict with our religious beliefs, in which science denies mystery, wonder and awe and replaced it with random chance. Random chance, it turns out, leaves us with a world that faces impossible odds, even given billions of years. Random chance was an idea that had to ‘stand in’ until we had a ‘better’ one. Now we seem to be developing a better, more explanative and complete one.
Creationists often attack the study of evolution as if it is attempting to destroy the miraculous nature of creation. I find that it does quite the opposite, replacing simple magics and children’s stories with one that is even more wondrous. I am not a Christian, but I do find plenty of room within the universe and creation for a ‘god-like’ presence. This is not a random universe, nor is it the one they taught us in Sunday-school. For me the world out there is entirely more miraculous and wonder-filled than that. The new stories out there, the New Phylogeny of plants, only add to our understanding and in so doing can teach us to be better ‘stewards’ of this Earth that we have inherited for the duration of our stay here. We are a conscious species and it can only benefit us to try to understand our place in this still evolving creation.
First Appearances and the Evolution of Plants
Life did not spring wholly formed on the Earth from ‘nothing’. The fossil record, our understanding of chemistry and the very nature of matter ‘says’ otherwise. There is a degree of complexity and constancy that supports the ‘idea’ of evolution. An ‘in-formed’, directed, universe. The universe and life demonstrate this and the odds of this being random chance are impossibly long. The relationships, contents and proportions required for everything between bits of matter to galaxies, are so precise, that something more ‘intentional’ is required to understand it. (Please see my previous posting, Life Inside the Cell: Waking Up to the Miracle, part 1)
I begin here with the formation of the Earth, 4,600 million years ago, or 4.6 billion (~4,600 Ma), roughly 200 million human generations or 46 million centuries, ago. Scientist calculate that the Earth first formed some 9.5 billion years after the creation of the universe. Most of us have trouble grasping even a single century. Our frame of reference, our own lives are hardly a speck on any timeline that we could draw and, because of this, puts such timescales beyond our ability to really comprehend them. Our tendency is to diminish prehistory and our own arrival which can make it difficult to understand the story of the Earth and its long and comprehensive transformations. Suffice it to say that much has occurred before us.
Some 500 million years, + or -, after Earth’s creation, came the first of the earliest life forms, primitive bacteria, prokaryotes, after the transformative chemical processes that created water. These first single celled forms of life which developed without a nucleus or the distinct organelles of the more ‘modern’ cells, eukaryotes, had their DNA simply held within the cell membrane with their other cell structures, reproducing by simple cell division, asexually. The first, Cyanobacteria, the only prokaryotes capable of producing oxygen, appeared more than a billion years later, beginning that long and essential process of creating an oxygenated atmosphere through an early form of photosynthesis, which most of life to follow depends upon. In fact, it is thought that this lead to the Great Oxygenation Event, some 2.4 billion years ago, when the atmospheric oxygen levels displaced methane creating aerobic conditions for the first time leading to a massive die off the previously dominant anaerobic bacteria and marked the beginning of one of the longest glacial ice periods on the Earth.
The story of plants on Earth is the story of life. Earth provided both the place and the raw materials necessary for its development and continuation. The sun still serves as the transformative energy that moves this process along while ameliorating the harsh, even toxic conditions that were present initially and continuing to do so even today. The availability of water, the precise levels of gases in our atmosphere, of O2 and CO2, the temperature ranges that characterize our various climates and regions, all go toward both determining what can grow here and, in turn, what those levels are in a huge complex system of feedback loops. As life on Earth evolved the growing conditions shifted as well, enabling ever more complex life forms to exist in a long string of necessary changes, each one dependent upon those that occurred before. Move anyone of these elements too far out of equilibrium and you threaten to disrupt the entire system. Move them fractionally and there is a strong conservative force that tends to keep the system in balance. Such changes over these many millions of years have resulted in a ‘system’ that grows ever more complex, a complexity that continues to provide alternatives for a changing world and the elasticity that helps ‘conserve’ the integrity of the whole. This same elasticity and complexity are built into the genetics of living organisms, beginning with these simple prokaryote cells, progressing up through the complex species alive today.

Overall, eukaryote plant cells, as illustrated on the left, are much more complex than those of the prokaryote shown on the right. The nucleus and the organelles are enclosed within their own protective membranes. The eukaryote shown is a more contemporary cell with its specific functions being met by more specialized organelles some of which are thought to be derived by including previously freely existing bacteria within their structures, i.e., mitochondria. Their complexity took millions of years to develop.
Early Cellular Life: First the Prokaryotes
The earliest cellular organisms, prokaryotes, were very simple, anaerobic, developing in water without the nucleus and several of the organelles, like mitochondria, that are characteristic of eukaryotes which appeared much later. What they had ‘floated’ freely within their cell’s cytoplasm composed of various lipids and proteins and primitive organelles that enabled life. Prokaryotes did contain DNA, a single strand, free within their membranes. Bacteria are an example. Again, their functions were ‘primitive’ accomplishing their metabolism, respiration, even reproduction following simpler ‘paths’.
Bacteria reproduce, asexually, from binary fission, simple cell division, not sexual reproduction. In bacteria as a result of certain internal ‘triggers’ a cell begins to double in size and doubles its DNA. After completing this stage the cell divides itself in half each with a precise duplicate of its DNA. There is no mixing of genes between individuals in the process. These early prokaryotes comprised a wide range of species that could occupy temperature ranges that would kill most organisms today, or specialized in more specific environments, which would have given them an advantage to occupying ancient oceans. The various molecules they might need were absorbed or even engulfed within themselves as bits of organic matter, alive or dead, by altering the form of their membranes and ‘growing’ around them. Cell walls are selectively ‘permeable’. There are thousands of bacteria today that can ‘eat’ and breakdown an immense range of organic compounds, in water. This is thought to have been characteristic of earlier bacteria as well and because of this they were instrumental in modifying the chemical conditions of their environments, converting compounds and building more complex molecules in the process, and including adding to the sequestration of carbon. Prokaryotes did not appear in a single prehistoric burst of speciation. New bacteria continue to form even today. There simplicity of structure remains and probably is part of the reason that so many bacteria are able to respond ‘quickly’ to changing conditions by forming new species influencing those environmental conditions. Again, it is their point of origin in evolution that dates them as the earliest cellular life form…not that today’s species and these first species are the same.
Prokaryotes rely upon the water that they live in, whether that water is ‘free’ or is within the structures of another organism, separated from it by their individual cell membranes. Sometimes cells merged, engulfed another bacterium, with specialized abilities, which radically changed the internal functioning of the cell improving their efficacy and thus their ability to succeed. This is widely considered to be how mitochondria came to be inside more modern cells. Maybe this was a bit like infection, maybe once in a several billion such occurrences, this gave a cell an advantage and this advantage lead to the dominance of its progeny? This may sound like random chance, but it is not. There is a progression, an advancement at work here, a genetic memory of sorts, a tendency to build upon previous successes and to shed what no longer works. Even regarding extinctions, there is no ‘retreat’ back to before, but a response that relatively quickly moves evolution ahead.
Water, in a very real sense, defines early cellular life. It provides the medium within which it lives and is essential for its life within its individual boundaries. If such a water dependent organism found itself dry on land, it would die, unable to meet either its internal immediate needs or to reproduce itself. This is still the case for many bacteria today as they may be unable to exist for a limited time exposed to aridity. A simple cell membrane may have organized and protected a cell within water, but it could not do so under dry conditions. These ‘simple’ structures and functions took many millions of years to evolve and have largely determined everything to follow.
Each additional step toward complexity has been important, many have been monumental and pivotal. This point is perhaps made more clear by the scales of time associated with each significant such ‘advancement’. In a sense organisms have ‘learned’ their ‘lessons’, manifested them in their structures and functions, retained them in their DNA, and ‘built’ on these over time, sometimes shedding what had proven useless or ineffective, when environmental changes no longer supported them via extinction. Physicists are beginning to argue that these lessons, these memories, are retained, outside of the individual organism, within the energy dense, super-fluid, quantum vacuum that fills the entirety of the universe (Please see my previous post, “Life Inside the Cell”, particularly the section titled, ‘Quantum Biology: The Realm of Quantum Physicis in the Organism’.) The more complex organisms became, the larger becomes their genetic ‘repertoire’ increasing their ability to adapt, to develop new ‘solutions’ to the problems they face. The basic structures and functions were well tested and established. The most primitive forms are still the most stable. The more evolved forms, of any given time period, with their more precise environmental requirements, are dependent on more complex relationships with other organisms and place, and, as such, are the most subject to failure and extinction. This leading edge of change, the present, is simultaneously the most diverse period and the least stable…and life continues to push this line ahead. Perturbations in environmental conditions have always been critical to this leading edge of evolution.
Eukaryotes and the Next Step in Evolution
Eukaryotes were the next major step in the evolution of life. Like their predecessors the prokaryotes, these too were water borne, but these contained a nucleus, the cell’s genetic material, its chromosomes, within its own membrane, setting the stage for meiosis, the splitting of the nucleus and its chromosomes in half, an early and essential stage in sexual reproduction. Sexual reproduction provided the way for the greatly increased flow of genetic material between individual organisms. Eukaryotes do not always follow this path. They can also increase through simple cell division, the process prokaryotes depend on exclusively. Simple cell division is still how Eukaryotes increase or replace the cells of the tissues of an organism. Among these Eukaryotes were the single celled species of Algae, that contained structures capable of what we now understand to be C3 Photosynthesis. This occurred around ~2,700 Ma. The way was then set for evolution of all plants to come.
It would take more than another 2 billion years to develop multi-cellular organisms, both plant, including more complex Algae, and animals, non-vascular, all still water borne – making their first appearance ~541 Ma. Multi-cellular life required that all an organism’s cells follow a coordinated and coherent plan, a degree of difficulty of then unprecedented complexity and magnitude. With their advent came the capacity for more complex and specialized structures within each organism. Before, each individual cell was relatively simple, limited and ‘complete’ in and of themselves. While floating around in the water they may have formed aggregates of individuals, but each one remained separate, limited in their capacity for complexity, though they certainly did diversify creating many thousands of species many of which have extant cousins today. With the ability to form multi-cellular organisms came the ability to build more complex ‘structures’ that increased the ability of these species to occupy different and more diverse niches and environments. It vastly increased their range of alternatives as life continued to evolve.
One of these more specialized cells were the ‘gametes’ that would become central to fertilization during the process of sexual reproduction, its task greatly complicated when one considers that within these two haploid cells were the entirety of the plans and operating systems for what would gradually become incredibly complex and impossibly large organisms. It is in the process of sexual reproduction, the creation of another individual, that multi-cellular organisms rely on meiosis, the production of gametes, by reducing their chromosomes in half, setting the stage for fertilization when the two would be rejoined in unique combination to form that which would become a new individual. Internal triggers begin this process, within specialized organelles of the cell, after an organism reaches maturity and the surrounding conditions, such as the season, would support the successful process. Once a new individual’s particular mix of DNA is created an organism follows it utilizing cell division to ‘build’ and maintain the organism.
Sexual reproduction was essential to an organism if it was going to be able to live on dry land. Organisms would have to develop tissues and structures that would allow it to do so. On land gametes could not ‘swim’ and survive. Structures were needed to guard it against aridity, not just to live there, but to make sexual reproduction even possible. The transport of gametes from one organism to another in early eukaryotes occurs in the surrounding water. While there is risk in this, the water provides a degree of protection. Sexual reproduction brought the genes of two unique individuals together greatly increasing the variability of the offspring, providing more ‘options’ for a species survival. This capacity was a keystone process.
Land Plants and Their Special Needs
Some botanists will argue that certain Algae were the first organisms to occupy dry land, at least those more marginal places subject to a regular cycle of inundation and drying, such as coastal areas under the influence of tides or somewhat longer cycles, as long as they were regular enough, for these early land organisms to survive. Timely inundation could support the movement of the male gametes, preventing their desiccation. But Bryophytes including Mosses, Lichen, Liverworts and Hornworts, all non-vascular plants, were the first fully terrestrial plants and they were and are, multi-cellular. Because they’re non-vascular they were limited by their inability to move nutrients and water between cells, limiting both their height and the distance anyone cell could be from water. (Hornworts are thought to be the closest relative to true vascular plants as they have intermediate internal, conducting structures) – These were the first sexually reproducing terrestrial organisms, appearing ~470 Ma.
There were several problems that had to be surmounted if multi-cellular plants were to be successful on land. Their permeable membranes that allows for the movement of water and nutrients relatively easily through it, could result in the desiccation and death of the organism on land. The ability to grow specialized cells and tissue enabled plants to form a protective epidermis and later a cuticle outside of it. These structures again took much time to evolve probably at a rate that enabled them to ‘move’ from wetter to progressively more arid environments. Adaptations were necessary to survive longer arid periods, drying out, slowing or stopping growth, until they could rehydrate themselves. Cellulose ‘happened’ giving plants the ability to better ‘armor’ themselves to aridity. Not having proper roots, these still had to rely upon taking moisture in through less specialized tissues.. Primitive root-like structures formed more for attachment purposes, which would allow them to remain fixed in more hospitable sites less subject to the stress of being ‘tossed’ about on dry land, but necessarily, always near water due to their lack of ‘reach’. Such primitive rhizoids, formed from different tissues than are found on the roots of Gymnosperms and Angiosperms, without the branching, hairs and various fungal structures that modern plants depend upon. These more primitive structures did not form the penetrating ‘networks’ within the soil roots do today.
It is also important to remember that soil did not magically appear either and it is essential if land plants are to establish and survive. Soil is a product of geological uplift, erosive and organic processes produced over vast periods of time and is directly effected by the organisms and living communities that live and die upon them. Arguably the life within the soil is as complex as the life above it. They evolved together in necessary stages. Decomposing organic matter adding their contents and various acids that would help ‘breakdown’ the rock and mineral soils into the complex structures of healthy soil, evolving into the dance of nutrients required for any healthy biome, building and holding what is necessary and valuable.


Ferns were the first vascular plants, appearing: ~360 Ma. They were, and still are, non-flowering, spore producing plants. Extant ferns, those species alive today, have all evolved from these first more primitive forms. The oldest species are long extinct their vascular and supporting structures evolving forming meristematic tissues that grew roots, rhizomes, stems and fronds, that contained the plant’s photosynthetic capacity and the location for the sporangia in which the plant produced spores, containing gametes, that could be released to the wider landscape. These early ferns were very simple, compared to the more modern species of today their fronds more modified stem than they were the more specialized leaf tissues they have today that are separate from stem tissues. Landscapes of this early period were largely swampy devoid of the ‘woody’ and flowering plants that we might associate with such areas today, because these all came many millions of years later. It was a very different world in both composition and appearance, a much ‘simpler’ world in terms of life and its complexity today…and this is a statement that could be made at any point in history.
It would be some 40 million year later when the first gymnosperms came into being. These include the conifers, cycads and the still surviving Ginkgo, considerably more evolved vascular plants with stems that produced ‘secondary’ growth, having stems that increase their girth over time from a cambial meristem. This capacity did not occur before. Gymnosperms were, and still are, non-flowering, (pollination takes place on a female strobili, a ‘cone’ like structure) seed producing plants, dating back to ~319 Ma. There were steps along the way before the arrival of the Gymnosperms, such as seed producing ferns, giant ancestors of our extant Horsetail species, Progymnosperms and their cousins, all now long extinct. Many of their traits are carried through to Gymnosperms today though somewhat more refined. Rare now, with their much diminished ranges, and threatened with extinction, is the entire family of Cycads. Again, during their earlier history, they came to dominate much of Earth’s landscape and are associated with an Earth whose climate had become drier, their root systems much more capable of penetrating soils more thoroughly and deeply.
It is interesting to note that the early development of stem structures lead early on to arborescent, tree like, plants…not the trees of today. The grasses that comprise a dominant share of prairies and savannahs, did not come about until the advent of Angiosperms. Early landscapes were more likely then to be forests in the sprawling swamps of the Carboniferous period. Lower plants were more likely to be relatives of ferns or early relatives of Horsetails and the many thousands of leafless, minimally branching, non-flowering plants.

Always thought that ground under your feet was solid? With the evolution of life the moving and splitting of land masses ‘explains’ much of links between species now long separated which also hugely effected the course of ocean currents which brought with it often radical swings in climate.
Between 394 and 299 million years ago the terrestrial flora evolved from one composed of small vascular and non-vascular plants to a vegetation that included ‘trees’, arborescent plants, towering as high as 100′, though none of these would be remotely recognizable to a person from today as a ‘tree’. This was a time of massive shifting of continental plates when the ‘super continent’ Pangea, and later, Gondwana, broke up and began to drift into their present, though still moving positions. There were massive changes and swings in climate. The poles themselves moved effecting the electro-magnetic radiation that struck the Earth. Shifts in oceanic currents with consequent changes in climate and weather. With the increase in plant life came a 90% drop in CO2 levels aided by the organic acids of plant growth, their roots releasing them to enable mineral uptake while converting silicate rock into carbonate minerals, as a side effect, that then moved by erosion collecting in strata on the oceans’ floors, sequestering massive amounts of carbon into the earth. Lignins developed in vascular plant tissues, that were more resistant to bacteria slowing the process of organic decomposition, resulting in what became the creation of coal and oil in the massive, warm lowland swamps that once covered much of the land, further decreasing carbon in the atmosphere. These same processes released massive amounts of O2, from the CO2 as Oxygen was freed from Carbon, into the atmosphere, increasing it from less than 1% to around 28-30% before it dropped to our current 21%. This later increase was aided by increasing levels of photosynthesis which releases free Oxygen as well. (During the period with the highest O2 levels there were ‘gigantic’ flying insects buoyed by the denser air, supported by the high Oxygen levels that could diffuse more deeply into their tissues. Insects don’t breathe, having no lungs, but still rely on their ability to absorb Oxygen to enable their metabolic processes.) Every molecule of biomass (CH2O) buried prevents its decomposition back to CO2 and H2O. This went on for millions of years. By extracting coal, oil and gas from these carbon ‘sinks’ and ‘burning’, oxidizing, them for their stored energy today, we are returning that now oxidized carbon back into the atmosphere. Dropping CO2 levels and the movements of the Earth’s land masses during this period lead to a cooling of the earth’s surface, and eventually to periods of glaciation and the extinctions that came with the temperature drops. With glaciation also came huge drops in ocean levels, by as much as 100 meters, and the exposure of the continental shelves. Overall, there has been a long term shift toward aridity, especially in interior portions of the continents. New land continues to form through volcanism and mountain ranges rise as continents continue to push one into the other and plates within continents grind into each other. The Andes pushed up some 15 million years ago radically changing climates on both sides strongly influencing growing conditions, that in South America, lead to speciation among the Bromeliaceae and others.
Stems and Stele: Developing the Strength to Stand
Stems are a radical development in the evolution of plants. With stems came a plant’s ability to add the dimensions of ‘thickness’ and height. Prior to this plants were limited to a more two-dimensional existence, living on the ‘surface’ or supported by the water within which they lived. Where the buoyancy and density of water once supported a few layers of plants in water, the atmosphere lacks these qualities and plants had to grow their own means of support, resistant to wind, that could support their top growth. Plants grow up reaching for the sun and with this ability create different levels of light intensity beneath and between them that shorter stemmed plants, with lower light intensity requirements can occupy. With stem development came a greatly increased capacity for plant diversity in a given landscape. Over time as stems grew taller they needed to be stronger and have more efficient vascular systems to support their internal functions. Stem strength was improved by increasing diameter and by ‘hardening’ the tissues. Some plants did this with a hardened ring of tissues near their outer circumference, others ‘wove’ a structure of stems together that transition to early primitive roots, not unlike present day Tree Ferns. The first of these early stem producing plants were relatively short and would have grown in low growing landscapes. Arborescent, tree like plants, came later.
The evolution of the stem is an example of specialization of structure, tissues and cells that began early on. Nothing like it existed before. It is thought to have evolved from more simple vascular tissues which are fluid conducting vessels within a plant, structures first evidenced within the Hornworts. Over time these vessels became longer, more continuous, adding cellulose and lignin, giving these vessels a degree of rigidity and durability to better and more efficiently transport fluids within an individual plant. Prior to this fluids could only move by capillary action between less specialized cells, ions and nutrients diffusing through tissues…not a very efficient method to meet the needs of an actively growing and metabolizing organism. Early stems greatly limited the height and thickness of a given plant.
Scientists study such seemingly inconsequential changes when able in the fossil record, and yes, they are sometimes found there. Then they compare them to similar still living species. Conclusions are drawn. Through the study of stems scientists have noted an evolution of their structures, again going from simple to more complex. Take a cross-section of any plant stem, examine it under a microscope and you can see what they are talking about, though to the casual observer, not looking so closely, they may appear the same. They contain different tissues and structures.
All stem forming plants begin with the basic stele. This is the central longitudinal structure running the length of the stem, tying it to the roots. Even our modern ‘woody’ plants begin this way, only later, over time these add ‘girth’ with the secondary growth of their cambium adding xylem tissue. Some stele have hollow cores, others are filled with spongy piths, some have their vascular systems in a more complete ring toward their exteriors, others are more diffuse scattered within their ‘cortex’. Some contain chloroplasts well inside this structure. Others create an exterior phloem layer which develops into bark. At some point the stems of some species began to form secondary xylem tissue, a process today recognizable in the woody Eudicots and many of the Gymnosperms of today. Others have more complex internal structures when looked at in section, sometimes almost ‘star-like’ in appearance, each indentation the location of a leaf base on the outside…all of these first form with a stele as they grow out after germination. Science has developed a timeline locating the originations these different structures each one originating after previous structures, each not necessarily completely supplanted by those that follow. These continue on today in various species…. This is true for many plant structures.
Leaves: From Green ‘Sticks’ and Stems

This is a 100 year old Welwitschia mirabilis growing in situ. These can live upwards of 1,000 years forming a ‘trunk’, which can attain diameters upwards of one meter, from which the reproductive structures form. The leaves grow continuously gradually drying and shredding at their end. This photo is from this site.
While the earliest land plants, mosses and liverworts, the Bryophytes, were photosynthetic organisms, they did not have leaves. Their tissues had not specialized to that degree. Stems came first and then a plant’s ‘ability’ to differentiate, to switch from ‘indeterminate’ stem growth and form the determinate, tissue of a leaf, ‘finish’ its growth and continue its stem growth. Leaves have a particular shape and size and, once complete, don’t add anymore growth (As with everything there are exceptions to this ‘rule’. See Welwitschia mirabilis a primitive plant of the Gnetales group and a Gymnosperm with the trait of growing two opposite, strap-like leaves, that grow continuously over the life of the plant…in other words, leaves with ‘indeterminate’ growth.). The earliest plants, did not form these specialized tissues, didn’t have the capacity to do so, and because of this had no leaves. This is also true of the earliest vascular plants, they grew only undifferentiated ‘stem’ tissue, a characteristic still exhibited in green stemmed plants today. Photosynthesis, as a process, occurred throughout most of these plant’s cells. This changed with specialization, as the thickness of plant structures increased and the CO2 levels of the atmosphere dropped. Extant plants like Equisetum spp., Ephedra viridis and Muehlenbeckia ephedroides, among many others, still possess this trait in that they include the chlorophyll rich chloroplasts near their stem surfaces. This trait is basic to even many, if not most, juvenile forms of plants, which are able to photosynthesize in the outer cells of their stems.
For the first 40 million years vascular land plant species had no leaves at all. CO2, essential for photosynthesis, was in much higher concentration in the atmosphere and because of this was able to diffuse into plant tissues more deeply and readily, supporting photosynthesis. The first ‘leaves’ to appear were ‘microphylls’ that were relatively small and were attached directly to the stem without petioles, not unlike the phyllodes of some species today, with ‘flattened’ stem tissue, still found on Acacia and other plants like Phyllocladus alpinus. Ancient microphylls, however, included only a single strand of unbranched vascular tissue. The Genus Seligenella possess microphylls and is still very common around the world, with species growing nearly to the poles with most occurring in wet tropical regions, and even a few desert dwellers.
Megaphylls, which developed later and independently, were more highly evolved from branch tissue and attached themselves to stems by a petiole, developed after CO2 levels and air temperatures had dropped to more modern and moderate levels. Leaf stomata, the openings in leaves that enhanced the uptake of CO2, were a later development as well, in step with dropping CO2 levels. These megaphylls later went on to become the leaves of modern Angiosperms. In virtually every case, plant structures and systems have been in a continuous state of evolution, as growing conditions have changed, inducing a change in the plants themselves, which in the aggregate often produce changes in the growing condition, all ‘regulated’, directed, by the possibilities contained within the DNA of plants and our very much evolving understanding of the A-field, the Zero Point Field or the Quantum Vacuum, which ever descriptive title you choose to attach to it. Over time a ‘record’ of these changes and possibilities accumulates…a puzzle systematic botanists have been working to unravel.

This chart shows the the approximate dates for the first appearance in the fossil record of these Angiosperm leaf shapes. Tha Aptian to Early Albian period was from 125 Ma to 112 Ma. The Mid to Late Albian was between 112 Ma and 100 Ma. The Early Cenomanian was the period just after ~100 Ma.
Angiosperm leaves evolved over this more modern period beginning with relatively simple, smaller leaves of an inch or so with smooth entire margins. The venation pattern was less dense as was the ‘density’ of stoma. They weren’t the efficient transpirers that they would later become. By roughly ~100 Ma, most of the modern forms were established including large palmately lobed leaves of early Platanaceae. These changes are thought to have given Angiosperms distinct advantages over earlier more ‘primitive’ plants.
Roots: Anchorage to Absorption of Water and Nutrients

Some Calamities specimen, members of the extinct Equisetophytes
Like other specialized structures roots evolved from primitive forms and were often similar to early ‘branching’ structures, and microphylls, modifications of simple vascular stem tissues. The fossil record shows first microphylls and later, branching, arising from nodes along the stem occurring typically in whirls. The were relatively short and simple without much differentiation in their structures and tissues. The Equisetophytes in the above diagram illustrate this typical pattern. Earlier plants exhibited even simpler structures with sparser branching. The above diagram also shows typical early rooting that exhibit a very similar pattern. These typically formed a running rhizome, a modified stem, from which stems would grow vertically from forming clonal colonies. These root/rhizome structures were thought to serve first as anchorage and secondarily for water and nutrient uptake. These plants tended to grow on wet swampy sites which dominated vast areas of early landscapes. This growth characteristic is still manifested in many of the Monocots today including rhizomatous grasses and bamboos in addition to the still extant species of Horsetails, Equisetum. These roots didn’t possess root hairs, nor did they branch. Root growth all initiated from these separated nodes of intercalary meristem in a way similar to the analogous growth on many Monocots today. They were relatively short and stubby.
Over many millions of years with the advent of Gymnosperms, the later Angiosperms and their ancestors, both stems and roots acquired the ability to produce secondary growth, adding caliper and the ability to branch providing better anchorage, their vascular systems developed improving a plant’s capacity to take up water and nutrients, allowing greater height and larger leaf surfaces. Many of these ‘woody’ plants evolved to form a spreading ‘mantle’ of roots at their base as well further stabilizing top growth. With their cambial meristem sheathing their roots plants became capable of forming ‘root hairs’ that were relatively short lived, with increased permeability and capacity to absorb water and nutrients. Root hairs were routinely shed adding material and complexity to the soil the plants grew within. While roots are largely beneath and beyond our ability to normally observe them they exist in a world every it as complex as the world above the soil line and in fact evolved in a coordinated manner with that upper, visible, world.
Sexual Reproduction in Plants: Sporangia, Strobili and Flowers
Over much of Earth’s history the oceans have provided a relatively stable, even beneficent, environment for life. One of these ‘benefits’ is water’s ability to serve as a medium for reproduction, both asexual and sexual, protecting and enabling the process in water, a moderating medium. Early sexual reproduction, didn’t occur until somewhere around ~1,200 Ma, and was, again, a simpler process than it is today in terms of splitting, matching and fusing of an organism’s gametes. By definition it still required fertilization, with genetic material ‘contributed’ by two independent organisms. The process of meiosis appears to have ‘borrowed’ from the earlier prokaryotes. Recall that they had a single strand of genetic material, a single chromosome, and that they reproduce by simple division of cell fission. They merely grow and then split evenly, each clone having an identical chromosome.
It is an extremely common occurrence that organisms suffer damage as they go about their business of living. When an organism is a single cell the likelihood that its chromosome could suffer damage is increased. Because these early organisms cloned themselves, chromosomal damage could be passed on to all of those that followed. Prokaryotes, however, developed a way to ‘repair’ such damage and thereby protect the integrity of the line, their lineage…and that involved a path very similar to the one found in early Eukaryotes and their process of meiosis. Prokaryotes would sometimes ‘engulf’ like organisms and, in a sense, there would be a kind of resonance which would activate an internal process that would ‘reconstruct’ their genetic material and thereby, the integrity of the line. Researchers now see this as a precursor to sexual reproduction and meiosis. This doubling of chromosomes, going from a single strand in the haploid cell of a gamete to the fertilized, diploid, cell of the zygote and new organism.
All of this took place within the protective limits of the cell itself and the surrounding water which would protect the gametes from desiccation. Other mechanisms and pathways had to evolve for this to happen on dry land. This pathway lead to the formation of a protective and encompassing spore which contained the gametes and later the seed forming plants of today. Before this was possible plants need to create internal vascular systems to move water, nutrients and metabolites around to their more specialized cells and tissues, to the structures necessary to fulfill the various requirements of the evolving plants.
Every plant and cell structure, all of the functions, developed over time with countless intermediate steps leading up to them. Like the Chicken and Egg story, there wasn’t suddenly a chicken that laid an egg. Seeds did not just spontaneously emerge one day from a flower out of nothing to fit our present and static concept of them. They evolved from simpler, more primitive structures, transforming from spores, tiny reproductive ‘bodies’, that formed in specialized tissues, sporangia, on tissues of plants. The sexual reproduction of multi-cellular plants go through consistent required series of stages. Non-seed forming plants came first, but the stages remain remarkably similar, flowering plants most recently..
Non-Seed Producing Plant Reproduction
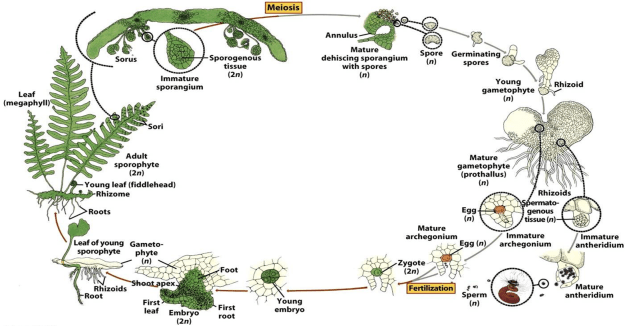
This is the life cycle of Pteridophytes, vascular, non-flowering, non-seed producing plants. Their life cycle is very similar to the more primitive, non-vascular bryophytes, the mosses, liverworts, etc…with two primary differences: the Bryophytes dominant, photosynthetic stage is their haploid, gametophyte stage producing separate male and female gametophytes which then produce the sperm and egg cells which upon fertilization produce the diploid sporophyte. In Pteridophytes, ferns, the diploid sporophyte is the dominant form we recognize and the gametophyte is much smaller, dioecious, containing both sexes, producing both eggs and sperm from different organs.
Male and female cells, sperm and egg, the genetic raw material, form within sporangia generally located on stems or leaves. These predate flowering plants. The sporangia came before the anthers and carpels of flowers, before the strobili, of Gymnosperms. The process of meiosis took place in these structures. The chromosomes are aligned and split in half, becoming ‘haploid’. Each half is enclosed within a spore’s protective shell. Spores, once released to water or the wind are carried away to germinate outside and independent of the ‘mother’ plant. After growing on, they produce either a male or female, intermediate organism, a gametophyte, from the gamete carried within the spore. In mosses this becomes the dominant form of the plant. In ferns, the gametophyte is small and the plant that we recognize as a fern is the second stage in the organism’s life, formed after the fertilization by an opposite sexed gametophyte. This forms the sporophyte, the seed producing form of the species. Their chromosomes fuse, doubling, forming the diploid form characteristic of all living Eukaryote cells other than the sexual, gametes. The above illustration is divided into a top, diploid half, and a lower, haploid, half. Sexual reproduction developed in a world that was warmer and wetter. Spores and the gametophytes they grow into, are more dependent on moisture putting them at a disadvantage when arid or dry periods occur. The male gametophyte, remember growing independently and separate from the female, produces sperm that must ‘swim’ to the female gametophyte in order to fertilize it. Enough water is absolutely essential. In many such plants the sperm may be carried in a splash or blowing droplet. Either way, water is required at these critical periods. (Here are two links to helpful videos. The first shows the life cycle of mosses and the second explains that of ferns.)
Seed Producing Plant Reproduction – Gymnosperms and Angiosperms

Welwitschia mirabilis a very early Gymnosperm forms these female ‘cones’ which the Welwitschia Bug pollinates. This picture comes from this site.
Seed producing plants first appeared around ~350 Ma. It took some 70 to 80 million years before they began to dominate the world’s flora. This was, again, another time of major climatic changes the most notable of which was increasing aridity, a condition that made it much more difficult for an external gametophyte stage to be successful. Many genera declined going extinct during this period. Cycads, part of the Gymnosperms, today are thought to have the most ancient lineage of all extant seed plants and can be traced back to ~318 Ma though, currently, extant species are of much younger derivation. At one time members of Cycadales could be found across much of the world’s landscapes, even on land approaching the poles which had much milder temperatures than today. As with other plant groups those that come to dominate often did so grandly their innovations and adaptations so well attuned to the climates and landscapes of the world. Later evolving groups and species in this process seem to run out of ‘steam’, perhaps having pushed the envelope of ‘type’ too far, the limitations of Cycadales now limiting their ability to dominate in a changed world. Today the 300 some surviving species can be found in only very small areas, many of them threatened or endangered. Other Gymnosperms, particularly the Conifers, are more widely represented around the world though the more recently evolved Angiosperms dominate them today in absolute numbers and by coverage of the Earth. Angiosperms, the Flowering Plants, still comprise the current leading edge of evolution a position they have held for the last 100 million years. Both of these seed producing groups of plants must go through the same critical stages to produce seed, but they do this in a slightly differently manner. (A short and simple video of the life cycle of a Gymnosperm.)
Neither Gymnosperms nor Angiosperms have an external and independent gametophyte stage. Instead this stage is reduced and contained, the male gametophyte, grows and ‘matures’ within the pollen including the formation of the pollen tube, connecting pollen to ovule, while the female gamete, grows and matures inside the ovule before fertilization. After meiosis, these cells increase minimally protected in their respective ‘shells’. Their gametophyte stage is very abbreviated and is protected from the aridity of the landscape.
In the later evolved Angiosperms the haploid gametes divide and double and double again inside the pollen and ovule, and the pollen is brought into contact with the stigma, by pollinator or wind, the remainder of the process taking place within the pistil and ovary, where it still remains protected from the dehydration that could plague it outside the organism. A pollen tube, initiated by one of the male gametes, grows down through the style into the ovary contacting the ovule. Through this move two more male gametes. They are delivered directly to the ovule. Once inside it one fertilizes the embryo, becoming a zygote, which contains the DNA and will grow into the embryo, while the other gamete fertilizes the two polar nuclei that have formed from two of the female gametes within the ovule before fertilization. These polar nuclei will grow into the endosperm, the two ‘seed leaves’, which will nourish the germinating plant that follows. Of course in Monocotyledonous plants, only one seed leaf is formed. The secondary, gametophyte stage then is entirely internal. The seed, the product of this process, contains its full compliment of chromosomes, in diploid or double form. It is protected within a seed coat and various dormancies that delay germination until such a time that growing conditions are more likely to support its growth while providing an initial supply of nutrient in the form of its contained endosperm.
Seeds, and the Angiosperms that produce them, are more complex in their structures and functions to accommodate these more evolved processes. By adding this complexity to their process Angiosperms reduce the hazards of aridity that they find themselves growing in.
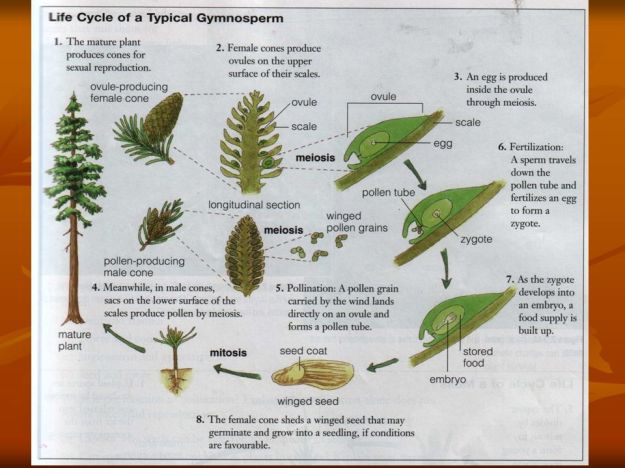
Life cycle of typical modern Pine species. Other extant gymnosperms as well as their extinct progenitors vary somewhat from this illustration.
The process of fertilization is somewhat simpler in the earlier evolved Gymnosperms in that they do not possess a true flower structure, specifically, the ‘female’ organs of a Gymnosperms strobili lack the ovary, stigma and style of a flower, nor do they possess anthers which release their pollen. These comparable structures in Gymnosperms are less evolved and simpler.. Male syrobili tend to produce a large amount of pollen, each grain possessing a pair air bladders, that cause the grain to drift on the breeze, increasing its likelihood of finding its female counterpart. The pollen is delivered generally by wind, but in some cases by insects, and the seed develops protected only by a ‘scale’, but is otherwise exposed or ‘naked’. Fertilization takes some time. Once the pollen contacts the ovule the ovule begins its processes of meiosis. This part of development is much slower than in Angiosperms. There is not an organ comparable to an Angiosperm’s ovary which develops into a protective fruit in tandem with the seed. There is then no fruit to attract fruit eating animals that will spread the seed. Just to confuse things some Gymnosperms do form with the immediately surrounding scales or bracts, fusing and swelling form a ‘fruit like’ structure that is attractive to certain animals that will consume it and later spread it by defecating. I know, I say to-may-toe you say to-mah-toe!






When visiting the San Francisco Botanical Garden in Golden Gate Park take the opportunity to stroll their Garden of Ancient Plants. There they have on display ‘relic’ plants, those species that have clung to existence, largely unchanged for more than 100 million years. Todays flora are highly evolved beyond these. It is interesting to note how some of these primitive structures are still being manifested in more contemporary species today, such as Cycads and Tree Ferns. Palms are still a very dominant group of plants in the world and their frond structures and the way they detach are very reminiscent of some of Earth’s first arborescent plants with their ‘megaphylls’ with multiple parallel veins that preceded modern leaves.
The First Arborescent Plants: Moving Toward Angiosperms
It should not come as a surprise that trees or arborescent plants came after much shorter and simpler plants. The oldest ‘tree-like’ structures found in the fossil record are Gilboa Trees, which grew to over 20′ tall, and have been found as fossils in New York state. They have been dated back to ~385 Ma. There were several forms of ‘tree’ that came and went after this date the earliest of which were all spore producing, all growing under swampy conditions. None of these formed ‘secondary’ wood, adding no growth to the girth of their trunks as is the case with most ‘trees’ today. Secondary wood, as we know it, is produced in many modern Eudicots, by their actively growing and dividing cambial tissues, which would not be around for a great many more millions of years. These were arborescent or ‘tree like’ plants that had a variety of internal stem structures. Think of present day Tree Ferns like Dicksonia spp. whose trunks are composed of a compact, twisting group of rhizomes from the base of which the roots grow out. Some of these had ‘buttressing’ roots developing above the soil and angling out and down where they anchored themselves. These early ‘trees’ evolved from leafless finely branching structures to those with the aforementioned microphylls and later to these with megaphylls, larger modified stem structures often divided into pinnae that would eventually dehisce, or fall away from the trunk or stem, much as extant Palm and Tree Fern fronds do, leaving large ‘scars’ on the trunks. The earliest Horsetails, Equisetophytes, with a structure that would feel familiar to us today, were among this group forming colonies of upright trunks, 4″-5″ thick, with one species, the Calamites, having grown as tall as 100′, from the spreading rhizomes below.
grew to over 20′ tall, and have been found as fossils in New York state. They have been dated back to ~385 Ma. There were several forms of ‘tree’ that came and went after this date the earliest of which were all spore producing, all growing under swampy conditions. None of these formed ‘secondary’ wood, adding no growth to the girth of their trunks as is the case with most ‘trees’ today. Secondary wood, as we know it, is produced in many modern Eudicots, by their actively growing and dividing cambial tissues, which would not be around for a great many more millions of years. These were arborescent or ‘tree like’ plants that had a variety of internal stem structures. Think of present day Tree Ferns like Dicksonia spp. whose trunks are composed of a compact, twisting group of rhizomes from the base of which the roots grow out. Some of these had ‘buttressing’ roots developing above the soil and angling out and down where they anchored themselves. These early ‘trees’ evolved from leafless finely branching structures to those with the aforementioned microphylls and later to these with megaphylls, larger modified stem structures often divided into pinnae that would eventually dehisce, or fall away from the trunk or stem, much as extant Palm and Tree Fern fronds do, leaving large ‘scars’ on the trunks. The earliest Horsetails, Equisetophytes, with a structure that would feel familiar to us today, were among this group forming colonies of upright trunks, 4″-5″ thick, with one species, the Calamites, having grown as tall as 100′, from the spreading rhizomes below.
The Broader Picture of Evolution
By the upper Permian (~260 Ma), the fossil record shows that seed plants began to dominate. Some 60% of plants were composed of Gymnosperms like Cycads, Ginkgo and other primitives like the long extinct seed forming Tree Ferns of the Glossopteridaceae, which once dominated the once super-continent of the southern hemisphere, Gondwana, throughout the Permian. Many claim that these Glossopterids were an important bridge species to the first Angiosperms as they share many details of stem and leaf structure as well. These other two extant genera are considered relics. The Ginkgoales, that contained as many as 18 different genera during this period are survived by today’s single last remaining species of the group. It’s a relic that contains many primitive structures that exist on no other remaining plant species. All of the other species of seed plants, that once dominated the earlier Carboniferous period, were in decline. The continents were continuing to shift, oceanic currents changing with them and the Carboniferous swamps that supported these early seed plants and the earlier spore producers, whose structures laid themselves down by the billions to be transformed into coal, disappeared as the Earth’s continents became not only more arid, but markedly warmer. The Permian was described as a period of change when the Earth moved from a heavily glaciated ‘ice house’ to an Earth as a ‘hot house’, completely free of ice. It is difficult for us today to understand how important this was. We think of the land and associated climates as fixed and unchanging, but it is anything but. Continents continue to move today at a pace beneath our notice.
Earlier scientists made links between plant species, genera and families and were able to piece the movement of continents, not just based upon their geology, but by the plants themselves, noting closely linked plants occurring on continents hundreds or thousands of miles away from each other. Genetic analysis has made this a much more precise process today. It is only with incredibly sensitive instruments that we can measure these movements and other phenomenon like land tides that daily cause the ‘solid’ earth to cycle, heaving itself up and down like the oceans, incremental shifts in the magnetic poles and changes in gravity itself. The ‘hard’ crust of the Earth floats atop a molten spinning planet pulled by the other massive bodies in our ‘neighborhood’. Sporadically, over Earth’s history ‘super-plumes’ have been created serving as slow flowing conduits of magma from deep within the Earth to its crustal surface, lifting the ocean floor adding significantly to the land surface, even forcing continents apart and into one another. Currents shift as bodies of water seek alternative routes in response to the differential heating by the sun, the repositioning land masses and the changing bottom and channels of the ocean floor. We build and live our lives as if the Earth is fixed….We are like fruit flies unable to grasp the scales of change in effect around us.
One of the most significant and recent changes to our living world involves the origin of Angiosperms, why did they appear so ‘late’ in the evolution of life here and how could they come to dominate the landscapes so quickly, other plant lineages disappearing along the way? These are unanswered questions still being addressed. We can look at this in a smaller way here briefly, by asking the same question of the ‘grasses’, the Poaceae.
Grasses first started showing up in the fossil record ~70-60 Ma. This is late as even most Angiosperm families had evolved prior to this. Currently there are upwards of 12,000 species including Bamboos and these dominate several landscape types, tropical and subtropical savannah, temperate grasslands and steppes, together covering more than 40% of Earth’s land surface. Huge such grasslands began to predominate some ~30 Ma. Obviously, prior to this there were none of these. Landscapes were mostly forested and early grasses first appeared as shade understory. This whole movement from forested biomes to grasslands as a result, at least in significant part, was due to increasing aridity and it occurred gradually and at different rates on different continents. Even today forested areas yield to grasslands as precipitation drops below certain critical levels. These is consistent with what we know about shifting continents and climates as continental interiors began to dry. We experience this today in our gardens when we seek to grow woodland plants when our own climates tend to be dry…we have to water them during critical growth periods.
The climates and continents continue to change as does the species composition of the communities and biomes that populate them. ‘Primitive’ species hang on in those climates and regions that still support them, that resemble their ancient landscapes. Overall growing conditions have been trending toward aridity and more modern plants have been trending in this direction becoming more resistant, while at the same time following an overall change toward more complex forms. New species have evolved and, in order to persist, they must both meet the challenges of their environment as well as the requirements of a living organism which must remain in balance as an individual organism. Whatever changes are ‘made’ must progress while ‘respecting’ the integrity of the organism, changes must move in concert with all of an organisms systems and environment. The increasing complexity of organisms over time emphasize the need/existence of the afore discussed ‘A-field’. Complexity and tolerances can’t progress by simple random chance.
As you familiarize yourself with ancient plants you begin to see ‘echoes’ of their structures in more modern plants. Because of the way genetics works, with the huge trove of code written into each plant, these ‘familiar’ forms keep repeating while internal ultrastructures and processes refine and make new places for themselves in ‘old’ forms. Because these structures can resemble primitive structures does not mean that they themselves are primitive. There is a continuous remixing of genes responding to contemporary conditions building toward a new equilibrium.
Evolution and speciation continues on today though one must look very carefully to see it, again time scales and the fact that our investigations are happening ‘now’, at the still leading edge of change. New species are likely forming, but it is very difficult to know which will establish and become successful, which ‘trend’ will win out. Extinctions are happening at a frightening and accelerating pace. Evolution has been a long slow process, one that has happened in bursts separated by periods of relative stasis…. I’m going to look for a moment now at how that might be.
Evolution is a ‘directional’ process, it is not random. Truly random processes don’t ‘advance’ or, for that matter, move back. Randomness implies the classic bell curve of scattered events. Evolution, however, is moving ‘forward’ toward more complexity, complexity of function and structure. It may not be easy for us to see at the leading edge of time, the always changing now, unless we can back away and examine larger swathes of time.
This is a screen capture from the Wikipedia page titled ‘The Evolution of Sexual Reproduction’.
Evolution, and the process of natural selection, was thought to be a very long and continual process, the result of various environmental stresses, many of which can be ‘read’ as corrections as I mention above. Conditions change putting species under stress. Species with traits that are unable to remain competitive in this changing environment, competing with other more adaptable species for a niche, would be at a disadvantage and would begin to decline. Certain individuals within a species might be more ‘favored’ because of variations within them that give them a ‘competitive’ advantage. Since every species contains a degree of genetic and physical variability this would slowly move a species into a more successful position…if possible. In some cases, populations of the same species might find themselves physically separate from others and ‘forces’ could, over time, lead what was one species to grow into two, now different, but genetically closely related.
Today most biologists agree that evolution is a long gradual process, but they also believe that it has occurred in bursts as a result of radical, even explosive, changes in the environment. Of course these ‘bursts’ are relative, in terms of geological time, not our own. Cataclysms, may present an opportunity for plants, in an appropriate position, to make a ‘leap’. For quite a number of years now many biologists have been arguing that something else could be at work pushing evolution that involved more significant and radical changes from what preceded them. If the environment provides the more ‘negative’ role, correcting ‘mistakes’, setting the conditions and limits for any given organism at a particular time. something else would seem to be providing the ‘impetus’ to moving evolution down its path of complexity. Some would attribute at least part of this to the wildcard of polyploidy, the doubling of chromosomes in some organisms that occurs most commonly when the reproductive process of meiosis does not take place. It is speculated that polyploidy can, at least some of the time, confer an advantage. I’ll look a little more closely at this in a later posting.
The Direction of Evolution and the Role of the A-Field
Evolution implies a direction, an evolution toward something. The study of the function of the cell suggests an ‘answer’. There is a ‘self organizing’ principle at work here. An intention or broader purpose even, that tilts the scales away from random chance towards complexity, diversity and coherence. Without this, life would be random, inchoate, it would never advance beyond some rudimentary form, its chances of success never over balancing those for death and entropy. Life’s first moments would never advance. The fact of evolution, the development of the cell, disproves this randomness. In fact it demonstrates the opposite. Life evolved in a now familiar progression and continues to do so though it is difficult for us to see this as our individual lives are so short, our senses limited and our record as a species brief as well. As corny as it may sound, there is a life force in operation here. Our reluctance to see it speaks more of our competitive, individualistic, even violent view of our universe than it does the actual world we live in. We choose to believe in a mechanistic world bound for an entropic fate, not its opposite, but this is observably incorrect. Such a world could have never produced such a complex and diverse living community here on Earth. It would have ‘eaten’ itself long before our world could have developed. We have mistakenly chosen to believe that life’s perpetual cycling, of reinventing itself, is somehow negative, even evil, as it ‘takes’ ourselves and loved ones. This is an incredibly myopic and immature world view. It is a refusal to accept that we are part of something greater. We insist that we are the pinnacle of life, God’s chosen in an otherwise expendable creation. In reality we are only relatively ‘recently’ arrived to this ‘party’.
The ‘cell’ is the success story here…proven and consistent. Multi-cellular organisms, those organized, and closely correlated, conglomerations of cells, communities if you like, have been very successful as a general idea and have spread around the planet, a planet itself that is not fixed in time, but is in a state of dynamic flux with, relatively short periods of stasis, geologically speaking, which allow particular organisms to dominate until changes again spur another marked change, building on what came before. The overall ‘direction’ of this evolution though is toward, as I said, complexity and diversity and has always been driven by a tendency toward cooperation at cellular, organism and community levels. The evidence is in the still evolving genetic record. Competition and death have always been a part of it, a limiting part of it, but not the driver. Limits alone would result in a dead world…and that is the model that has been dominating our world view lately.
The cells of organisms are composed of molecules which are composed of ‘quanta’ the energized, vibrating strings of modern string theory. They resonate to various frequencies within the Zero Point Field, ZPF, depending on their their structures, their species. As long as they are alive they resonate at specific ‘frequency’ and they contribute to the field itself. Upon death the organism begins to break down, decomposing into its constituent parts, available to other organisms, responding to other resonances while the field continues, ‘remembers’ and moves ahead, supportive and directional, cycling. (Please reference Laszlo’s book below.) Death itself ensures that this process of evolution continues. Immortality, would stand in its way.
The ZPF, the Akashic or ‘A’-field, the quantum vacuum, which ever label you select to attach to it, ‘in-forms’, influences everything. It does not determine it. This is where cutting edge physics theory aligns with biology. The universe does not have a specific ‘goal’ in ‘mind’…it does have a consistent ‘tendency’. Ervin Laszlo, the widely recognized and celebrated, scholar in ‘systems theory’ and the philosophy of science, describes its ‘behavior’ as a kind of ‘game’ of 20 questions, in which possibility and creation continuously ‘ask’ questions and the A-field responds back with consistent ‘answers’, subtly directing the course of ‘play’, but not the ultimate outcome. The A-field is in continuous direct, coherent and harmonic contact with all things, all organisms, a conserving influence, while it takes a guiding role in the process of evolution. This direction is proving to be demonstrable over the vast stretches of the Earth’s and the universe’s existence. Life, complexity, coherence, correlation, are all consistent themes within the A-field. Death is merely part of the cycle, one that can prove dangerous when too much is made of it, when one species moves out of balance with the rest..
Conclusion
Life is not static. It can be defined by its dynamism. Evolution has been both a steady and ‘explosive’ process and we should expect that it continues to be, including massive extinction events. Today’s extant species are not an ultimate end point, they are only our present position, a moment in time we call ‘now’, the result of changing conditions, extinctions and speciation, both rapid and gradual. The occurrences of polyploidy helps to emphasizes the role of genetics in life and evolution. The work of systematic botanists and the APG shines a light on all of this.
Evolution ‘occurs’ at the leading edge of time, running in a continuous wave, including every species, every element and force…every moment. This is where life happens, procreation, consumption, maturation…and death, speciation and extinction, every species, a piece of it, subject to it, as the conditions change and cycle themselves. Evolution moves in the direction of increasing complexity, at both the level of the species and as an overall community of life. It is not the individual that matters, not necessarily even a particular species, but this overall increase in complexity and diversity. In the process species are added..and they are lost. Some survive and continue to evolve over the course of many millions of years while many others serve a much shorter term purpose. The simple bacteria of the prokaryote group, the first successful form of cellular life, are supremely adapted to the changing conditions while they undergo a continuous process of speciation. Other species have endured for many millions of years, no where as long, but a remarkable feat, while countless thousands of species have been here and faded away…all of this happening here! now! Success does not have some ultimate measure, it is what moves life ‘ahead’. Maybe what we really need to learn are new stories, stories that we can access, that can take the gleanings of this new and evolving science and tell it to us.
If we as a species are going to continue here, understanding this process has become essential. It is in understanding the intimate relationships between the many thousands of species that we can begin to unravel and appreciate the wondrous world in which we live. To continue to insist that the world is a fixed creation, devolving in a competitive ‘orgy’ of consumption and violence, to an ultimately entropic end, is to ignore what life essentially is. Our ‘societal’ rejection of even the idea of evolution is a result of fear and ignorance, a blind insistence that we are in control of what matters in this world, or that God created all of this just for us to destroy in a demonstration of our own weakness. This is a rejection of the world and is both naive and dangerous, an act of ultimate hubris, that will eliminate us from the unfolding story. This science may seem to be a pointless esoteric exercise to many, but what systematic botanists and the many other scientists at work within the APG scientific community are doing here, is necessary knowledge as we as a species continue to increase our numbers and our impact upon the living Earth. It has become a matter of our own survival as a species, that we understand this and change they way that we live here.
Systematic botanists are helping to unravel this mystery, drawing links and making it possible to suggest how plants and life may have evolved to today’s level of complexity and diversity. Their work is giving us a new appreciation of the value of all of the life here, the product of several billions of years of evolution. Understanding where we came from and how we got here is becoming ever more important to our own survival. When we come to better understand the integrated and coherent forces that determine the health and vitality of the planet upon which we all live, we can become true stewards of this life and gain a little of the humility that we as a ‘conscious’ species have chosen to ignore. As a part of nature our actions as individuals and societies, play a powerful roll in shaping the conditions on which every species depends. We are entirely capable of extinguishing whole species, and have in fact done that many times over, just as we are of selecting others to preserve and protect. Our challenge is to understand and define our role as a member and participant in this miraculous experiment of life. With our numbers, technology, economy and political choices we have chosen a path in opposition to the creative aspect of life. We have seized the role in this game of 20 questions and have been all too consistently answering its questions violently, ignoring the several billion years of history and evolution that got us here. People often scoff, dismissive of now extinct species, particularly of dinosaurs, that were essentially too ‘stupid’ to survive, forgetting the millions of years that their line inhabited the Earth. In comparison our existence threatens to be but a brief catastrophic moment….
[I will be adding a fourth post covering the role of polyploidy in evolution. I promise it will be substantially shorter. Of all of these it is the ‘newest’ topic for me.]
A Short Bibliography
The Evolution of Plants 2nd ed., K.J. Willis and J.C. McElwain, Oxford University Press 2014. This is a very accessible introductory college level textbook on plant evolution which introduces much of what botanists, geologists, etc consider when investigating the phylogenies of plants and their evolution along with the atmosphere, soils, climates, shifting continents, water bodies and the many structures and processes within plants themselves. They discuss evidence from the fossil record and genetics as well as the morphology of plants today. It avoids an in-depth treatment of plant genetics. I found this book to be very helpful without being too ponderous.
‘Following the Vascular Trail: The Path of Water from Soil to Atmosphere’. This is a previous post of mine on the movement of water within the plant and the three photosynthetic pathways which evolved over time to meet the variable climatic conditions across the Earth’s landscapes.
The Fourth Phase of Water: Beyond Solid, Liquid and Vapor, Gerald Pollack, Ebner and Sons, 2013. This is Pollack’s second and more thorough look at the magical substance and capacities of water by the University of Washington bioengineer with a gift for putting the technical into very accessible layman’s terms. It is still not a quick read, not because of its jargon..but because there is simply so much that is new for us to grasp. There is much here that speaks to the powering of the organism and life.
I include this simply as an example, “Ginkgo biloba – Its Ancestors and Allies”, Zhiyan Zhou, a page on the International Organization of Paleobotany’s website. Such ‘tastes’ help us gain some perspective on the enormity of the undertaking facing botanists and taxonomists in their work.
“Morphological evolution in land plants: new designs with old genes“,
“Origins of Eukaryotic Sexual Reproduction”, Ursula Goodenough and Joseph Heitman, Cold Spring Harbor Perspectives in Biology. This is a relatively current review of the scientific literature covering the topic and while quite detailed, and extensively footnoted, is not so technical that the interested layperson will get lost in it. I read it twice, scanning quickly for the broader picture without getting hung up on details beyond me and then more carefully once I had an understanding of what it had found. The jargon is fairly technical, but depending on your background, as in my case, I know enough to read ‘around’ the rest.
The Rainbow and the Worm, 3rd. ed., Mae Wan-Ho, World Scientific Pub., 2008. In this volume Wan-Ho discusses the quantum nature of the cell and the organic architecture of living organisms, the bio-chemical and electrical functioning of an organism. Over the last 15+ years I’ve read each successive edition and glean something new from it each time. Don’t pressure yourself to get it all on the first go….give it some time. Well worth it! There is a later title, Living Rainbow H2O, by Wan-Ho from the same publisher in 2012, that looks even more deeply into the essential role of water…recommended!
Science and the Akashic Field: An Integral Theory of Everything, Ervin Laszlo, Inner Traditions, 2004. Laszlo is a former professor of philosophy, systems theory and futures studies, the author of 74 books and over 400 papers. This book presents his interpretation of the Akashic Field, which is equivalent to science’s ‘zero-point field that underlies all of creation. It is the constant and enduring ‘memory’ of the universe containing all that has ever happened, preparing the way for what is to come. It is the lynch pin in all of this.
Unflattening, Robert Sousanis, Harvard University Press, 2015. Sousanis is Assistant Professor of Humanities and Liberal Studies at San Francisco State University. He majored in mathematics and went on to become the firsts doctoral candidate at Columbia University to write his entire doctoral dissertation in graphic novel form. His dissertation was later published as this book. There is a whole movement, if you will of, scientists/artists, beginning to reach out to the public through comics, a graphic form, to present what has previously been inaccessible to the interested lay person. Here is another such title, an effort which Sousanis participated, “Eating in the Anthropocene” in which participating comics creators translated specific ideas around biology, geology, agriculture and food security moving into a future of climate change and population growth developed in a symposium in Europe where comics are more widely accepted across age groups. The link that follows presents all of the formal papers presented there and the idea. “Science Meets Comics. Proceedings of the Symposium on Communicating and Designing the Future of Food in the Anthropocene”.science-meets-comics-ebook

This was a very interesting read! I particularly enjoyed the part about structure modification (growing a stem to attain height, etc.)
Thanks for taking the time to share the information.
LikeLike