

I don’t know what this Bromeliad is, but it is statuesque, the inflorescence reaching well above my head. I took this shot next to Burl’s ‘chateau’ at Rare Plant Research just south of Oregon City. He moves a lot of tropical exotics in and out of his greenhouses every year. This is what a lot of people think of when they picture a Bromeliad.
I awaken and come down stairs at about 7:00 am…it’s a warm 66ºF outside. I was up late last night, until after 12:00 am, keeping the air flowing through downstairs in an attempt to cool the house. This is on the warm side for us here in the summer. On rare occasions our lows can drop to as high as the low 70’s…such temps tend to occur more frequently in more recent years when ‘heat lows’ settle in around us and we suffer through ‘heat alerts’, whenthe air stagnates and turns ‘brown’ and we can become caught in one of those cycles of days where our highs remain in the upper 90’s and low 100’s. Our all time record high of 107º, in August of 1981, was during such a cycle that I had the privilege of experiencing as I was here in Portland visiting a friend and attending my brother’s wedding. On the 6th it hit 99º. The high rose the next day to 102º, 105º on the 8th, 104º the next, 107º on the 10th, the humidity at 15%, then cooling to 97º on the 11th. I remember taking turns trying to cool ourselves, without any air conditioning, submerging in a tub of tepid bath water,
As our climate warms such temperatures are becoming more common. In June of ’15 we recorded 9 days with highs of 90º and above, 21 over 80º, the warmest June on record, a month that has historically been a ‘spring’ month in terms of its variability. That month saw several other records fall: consecutive days without rain, previously 18 now 25, and warmest June night, once 68º now 71º.
[We are currently, as of July 29, ‘working’ on day 42 with zero precipitation…and none in sight! As most gardeners know this does not ‘balance out’ our heavier than normal accumulation thus far this rain year. Plants need water while they’re in active growth, not dormant. We’ve hit 100ºF once, it was 102º on June 25, had 3 days at 90º or better and 8 over 80º, a fete we almost duplicated in May this year. In July we are shaping up to be quite normal averaging around 2 degrees F over our normal high of 81º. The forecast, however, is alarming. We are projected to have two days in the next week that could break our all time high record, the forecast calling for 109ºF. Climate change is not about today or tomorrow, its the big picture, the extremes and patterns, the changes to the margins, what we and what lives here has to stretch to accommodate.]
It has not been uncommon historically to have many June days locked into a cool drizzly pattern with highs stuck in the ’50’s, the maritime influence still strong and retaining its hold over the daily weather. Such cool/wet June’s are so frequent here that there is a commonly used term for such a pattern, ‘June-uary’! a pattern that is remarkably like many of our actual January days, such can be the effect of the cold north Pacific 90 miles to our west. People used to joke about how summer in Portland begins on the fourth of July. We are warming.

These two graphs of the fateful, for my Fascicularia pitcairnifolia, winter of ’13-’14. Showing our typical, erratic, spiking pattern of temperatures with two abnormally cold periods. The lower graph shows the years accumulated percipitation, 5″ below ‘normal’. In contrast, through July of our current rain year, we have received around 57″ in inner SE Portland, about 23″ over normal, or 67% above normal. The time period is our ‘rain year’ measured from Oct.1 – Sept.30 beginning in the fall after our typical summer ‘drought’ period.
This is probably part of the reason that i’ve found myself drawn to plants from historically warmer climates. Experiencing increasingly warm weather days, many once well adapted native species can find the climate ‘welcome mat’ less hospitable than it has previously been. Shifts in average temperature, higher peak temps, have a cumulative effect on living communities in which little shifts can potentially lead to the loss of marginal populations which can in turn have a greater or lesser effect on other species who have depended on these for food or had symbiotic or commensal relationships with them. All relationships are complex. Growing plants from other climes and regions gets you looking more at why there and not here. What kind of changes are necessary for new, previously exotic species, to find a new home here filling niches left by those not tolerant of our changing conditions. Climate change aside there are still the hugely altered growing conditions of the city, its heat island and drastically compromised soils, the introduction of so many weedy and invasive species and the surrounding regional landscapes, either put directly to human use as agricultural lands, developed more directly for human occupation…or simply heavily disturbed ‘transition’ lands, serving no direct purpose, but forever changed. I write of this more directly elsewhere.
What fairer weather plants have I been thinking about? family, Bromeliaceae, a plant family, containing 51 genera and around 3,500 species, none of which are represented here in our native flora, one of the last families most gardeners might think about when considering plants for the Pacific Northwest. Typically these are thought of by most of us living in temperate climates as ‘wet’ tropicals, but the family has proven adaptable and includes species that can grow well beyond the tropical and sub-tropical regions of the world to where they’ve been introduced into Mediterranean regions, proliferating in many California gardens as far north as the Berkley area. I grow one species, in a ‘protected’ spot, in the ground, in my own garden.
_________________________________
A Few Words On Vegetative Armament, Plant Defense and Recurved Spines: Jaws that Bite, Claws that Tear
I spent almost 30 years working in Parks here, in the ‘field’. My tasks were many and aligned with the landscapes and properties under my charge. Invariably, much of my time was spent with the control of weeds and invasive plants. In particular I want to talk a little about Himalayan Blackberry, a Rubus sp. that has been assigned several species names over the years, most recently Rubus armeniacus, that I learned as R. discolor. I worked in north and northeast Portland and later on in Parks in Downtown and the West Hills. In some of these parks this plant covered many thousands of square feet, if not acres. I battled it at Kelley Point and Pier Parks where it occurs in large patches, in forest openings in Washington Park and stands of it on the steep bank below the Eastbank Esplanade and the west bank below the Riverplace Esplanade where it constantly tried to spread into more desirable vegetation. The conditions at each site shaped my strategy in a way I thought would be most effective and safe, both for me and the surrounding landscape.
I can remember many times ‘wading’ into a mature stand of Blackberry 8’-15’ tall, covering several thousand sq.ft. in a ‘patch’, with my gas powered hedge trimmer, slicing and dicing, as I moved along its edge and ventured into its mass. It was a workout to say the least, cutting with broad strokes low, medium and high, horizontally, vertically and diagonally, left and right. If the area was very large, this was a winter job, heavily protected in coveralls, chaps, gloves, boots, a helmet and face shield in an attempt to armor myself against the plant’s defenses.
(Later, in late Spring or early Summer, I would return to apply a broad-leaf selective herbicide on the much lower and more compact regrowth, reducing the danger of its drifting off target, damaging non-target species and making it safer for me.)
The tactics involved cutting it as I went, into manageable pieces, that would drop where they were cut without undermining the ‘structure’ in such a way that the whole thing might ‘collapse’ on top of me, each reverse curving barb perfectly positioned to bite into clothing or my skin if presented. Of course this would happen anyway.
Other times, when the particular ‘incursion’ was much smaller or growing within desirable plants, I might press my way in to reach the bramble’s base, or where it may have tip layered, and cut the stem back close, if I couldn’t pull or easily dig it out, immediately treating it with a concentrate, using a squirt bottle, minimizing the chemical used in a directed and precise way. This method could bring you into close contact as well with the business end of the plant’s barbs.
Some plants are merely spiny…they stab when you get to close and withdraw as you back away, not this particular Rubus. You learn very quickly that once ‘engaged’ by the Blackberry, a quick and hasty retreat is a mistake, as the recurved ‘claws’ of the beast sink into the soft tissue behind your ear, your neck, arm or leg. What is required is calm. The claws of big cats are recurved to help them hold their prey preventing escape. The effect in the vegetable world is similar. So, think! You set down the hedge trimmer after shutting it off. Evaluate the severity of the attack, the critical points of contact, while still engaged, and then thoughtfully and dispassionately, move the aggrieved limb or flesh, forward, into the mass of the plant, to release the barb’s hold while lowering, twisting or lifting the offending cane away…moving your body in the opposite direction once the claw’s hold is released. This action is complicated when there are several simultaneous points of contact in front of you, to your side and even behind you. When cutting, it is absolutely essential that you anticipate what effect your cut will have on the mass of bramble above you, how and in which direction it will collapse. Blood will be lost. If there are too many points of contact, attack, it can get ugly in a hurry.
Recurved spines are not unique to Blackberries…fortunately they are less common than the straight spines so common to many other species at least among the plants we use in horticulture. They occur along the leaf margins of many Agave spp. and are very common on many members of the Bromeliad family. Interestingly, those plants we commonly call ‘carnivorous’ the Sarracenia spp. and Darlingtonia, don’t have these, they pursue an alternative strategy. Some of the Bromeliads can be merciless with anyone or thing that gets too close. It wasn’t that long ago that I didn’t grow any of these plants, I’d even eliminated Roses from my own garden though that decision had nothing to do with their thorns. Among these plants, Agaves came first for me.







Their attraction was architectural and my curiosity was piqued by the whole desert and monsoonal rain pattern thing. Before too long I had begun adding members of the Bromeliad family, staying away from Cactus as my limited knowledge and experience with Opuntia and Cylindropuntia, two genera renowned for their ‘glochids’, the tiny hairlike, barbed spines at the base of their more obvious spines. The glochids could ‘bite’ and keep biting, from seemingly incidental contact, and they seem to do this even when the gardener, or visitor, has avoided contact with the much more visible projecting spines. The ‘spineless’ Opuntia‘s still have the troublesome glochids giving the ignorant an unearned feeling of safety. At this date I have slightly relaxed my Cactus ‘ban’ and have tried a couple of the ‘fish-hook’ type cactus, which are glochid free and have ‘bent’ tips at the ends of their spines reducing the incidence of casual punctures.
Botanist often argue that these various kinds and sizes of spines, of ‘armament’, are defensive, protecting the plants from the many grazers and browsers…but, I have to ask, is that all it is? Why thorns, sharp little bristles, those irritating little hairs that some plants have and…why glochids. God, why glochids? Why not just toxins and chemical irritants? Is it really only to deter ungulates and browsers? Are these vicious hooked barbs the only thing that have saved so many Bromeliads from oblivion? Are they otherwise so darn tasty that all of the grazers, gnawers and chewers would decimate them and leave us only with grasses and sweet soft herbaceous plants? I doubt it. Why haven’t any grasses developed such defensive barbs? True, grasses have the ability to ‘bounce back’ from normal grazing ‘pressure’ and the are prodigious seeders and many, especially when ‘cropped’ down, spread move thickly, vegetatively, across the ground. What about all of those woody species without any ‘weaponry’ whatsoever to defend themselves and future generations? that are slow to reproduce? and, when they do, do so sparingly? Are these little ‘eruptions’ and exclamation points along a Bromeliad and Agave leaf’s margin just some little quirk of botanical fate? Were the spines of Rubus spp. really ‘created’ by the relentless chomping of herbivores? Will Hawaii’s thornless Rubus spp. develop similar spines if the exotic goats now there are left on their own to munch what they will? Is such spininess more common and aggressive on species that take years to reach maturity? If I did a survey would it show that slow growing plants, plants that never grow beyond the reach of local ‘predatory’ herbivores, are more heavily defended than others? Are most plants that utilize the CAM photosynthesis pathway, which contributes to some plants being slow growing, likewise heavily defended? Bromeliads are such CAM plants, so are Agave. How have the grasses and sweet clovers survived so long? Are they just god’s chosen sacrificial children? able to recover from regular attack? Do these barbed and spined plants have some inherent value that the world can’t simply live without? So many questions.

This still emerging Eryngium agavifolium, at this stage, looks more like it could be some kind of Bromeliad than an Agave. Though it looks vicious you and your baby soft skin are much safer around it than similarly barbed Bromeliads.
Whatever the reason for their existence, such marginal armament, is both a deterrent and an attractant to gardeners. The architectural forms of many of these arcing, rosette growing plants, appeals to many of us. Their spines, straight and recurved, serve as repeated decorative motifs that we find beautiful, natural creations, precise and perfect, that would take skill and concentration for us to create on our own. Such plants also ‘demand’ our respect and do not ‘suffer fools gladly!’ These are qualities many of us admire.
For us gardening in the maritime Pacific Northwest, desert, tropical and sub-tropical plants tend not to be plants for novices. They are more of an acquired taste here, that come with maturity, experience, I like to think, or…I suppose it could be a very specific type of ‘dementia’ some gardeners can be subject to. Why does an otherwise intelligent, sane, person, start growing plants that can stab, rake and otherwise bloody them, just as their skin is beginning to thin? Despite our best efforts our skin becomes more fragile, thinning, and much more subject to damage as we age. Now, when I lift a pot of Hechtia ‘Texas Red’ to set in a tray to bottom water it, it is all too easy to inflict little tears to the backs of my hands and forearms because of the stiff barbed foliage that reaches down near the pot’s sides. I divided a similarly structured Dyckia ‘Red Devil’ a few years ago…there is no give in these plants and each has the ability to grab and hold. Just getting the stiffer of some of these out of their pot is a challenge. It would seem that Bromeliaceae, have evolved their structure to frustrate and fend off would be gardeners, at least those species that are stiffer leaved and more aggressively barbed.
Bromeliaceae: Welcome to the Family
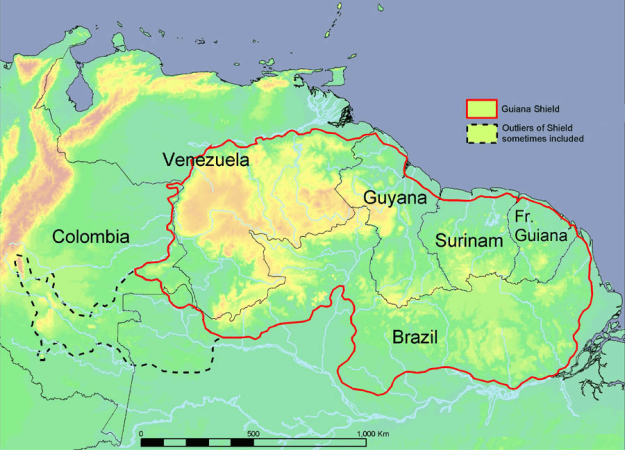 Today’s Bromeliads can be traced genetically back 70 million years to the Guiana Shield of South America, a ‘craton’, composed of incredibly dense base rock material from the Earth’s mantle, heaved up through the crust, which is, even in geological terms, very slow to erode. South of this craton, is a huge basin, that has twice been a huge inland sea, some will argue whether it was freshwater or salt, which has become, for the last several million years at least, the Amazon Basin containing the huge river of the same name. Three of these cratons comprise much of the continent of South America one of which defines the Basin’s southern border, underlying the Brazil Highlands. The Guiana Shield provides the ‘foundation’ for most of the northern coast of South America, extending westerly from French Guiana, across Surinam, Guyana and most of Venezuela including a portion of northern Brazil. Sometimes geologists include part of Colombia. This area was a lowland tropical jungle, with similar conditions to the present day Amazon basin, where early Bromeliads flourished as terrestrial C3 plants. The Shield had not yet pushed up to form the Venezuelan Highlands further dividing the continents of North and South America.
Today’s Bromeliads can be traced genetically back 70 million years to the Guiana Shield of South America, a ‘craton’, composed of incredibly dense base rock material from the Earth’s mantle, heaved up through the crust, which is, even in geological terms, very slow to erode. South of this craton, is a huge basin, that has twice been a huge inland sea, some will argue whether it was freshwater or salt, which has become, for the last several million years at least, the Amazon Basin containing the huge river of the same name. Three of these cratons comprise much of the continent of South America one of which defines the Basin’s southern border, underlying the Brazil Highlands. The Guiana Shield provides the ‘foundation’ for most of the northern coast of South America, extending westerly from French Guiana, across Surinam, Guyana and most of Venezuela including a portion of northern Brazil. Sometimes geologists include part of Colombia. This area was a lowland tropical jungle, with similar conditions to the present day Amazon basin, where early Bromeliads flourished as terrestrial C3 plants. The Shield had not yet pushed up to form the Venezuelan Highlands further dividing the continents of North and South America.
This separation of North from South America and the uplift of the Guiana Shield set the stage for the development of today’s Bromeliads. Genetic studies show, thus far, that the family was quite stable for some 51 million years, up until around the time that the Andes began to push up, (there is no agreement as to when the Andean uplift occurred, different camps arguing variously for dates from 15 – 30 million years ago.) closing the last lower elevation avenue for genetic material to move north and the time that the Amazon established its current form. This effectively divided the Family from any more intermixing. What was to the north would stay and stabilize, changing independently to the conditions extant while the genetic material to the south responded in it own way to its varied terrain and conditions. The land’s of the Shield transformed drying as they heaved upward. The Andes became a major factor in the continent’s climate, containing much of the precipitation, brought in by the predominant trade winds from the Atlantic, to the huge basin and the eastern side of the mountains while simultaneously creating an extensive ‘band’ of aridity along much of the Andes’ western flank and interior. (It is also believed that prior to the Andean uplift that the Amazon flowed westerly to the Pacific!)
Bromeliads began invading drier areas in Central and South America roughly 15 million years ago. The group underwent a process of major adaptive radiation involving the repeated evolution of epiphytism, CAM photosynthesis, impounding leaves, several features of leaf and trichome anatomy, an accelerated rate of ‘speciation’ and the creation of new genera. Today there are arguably eight sub-families, an expansion beyond the former 3, based on morphological studies alone, according to botanists who study the Family. Two of the original sub-families Bromelioideae and Tillandsioideae remain, largely unchanged, as they have proven to be monophyletic with a linked ancestry. Pitacairnioideae, however, was determined to be paraphyletic, being more of a genetic hodge-podge and so was broken down into 6 other sub-families.
Much of the process of speciation, that began in earnest 15 million years ago, was through the process of adaptive radiation, an evolutionary biology term for those periods when significant environmental changes occurred making new resources available, creating new challenges, or opening new environmental niches. Species respond to these changes exhibiting physical, morphological, changes, often to the degree of forming entirely new species. As a group the family began to shift from being terrestrial plants, rooted and growing in the soil, to epiphytes, attached to and dwelling on trees. Over 1,800 species in the family are epiphytes, using their roots to anchor themselves to other plants, without parasitizing the supportive host plant, gathering their water and nutrients via their above ground structures from the water and decay around them. Many others are lithophytes, anchoring themselves to rock. One way that Bromeliaceae have evolved to these environments is by altering their basic rosette form into one with impounding leaves, their leaves emerge, without petioles, tightly overlapping, forming a ‘cup’ in the center of each rosette in many species, sometimes substantial in size. This structure ‘funnels’ water that strikes their top surface down into the crown, in many cases capturing and holding it. This structure is less pronounced in most terrestrial Bromeliads which instead funnel the collected water to their absorbent roots. Many epiphytes do the same with fallen organic matter. These are self limiting structures capable of holding only so much depending largely on regular rain and/or cloud condensate to meet their needs. If there is not enough on a frequent schedule these plants will decline and possibly die.
Many of these epiphytic Bromeliads, like hundreds of other epiphytes, including many Orchids, trap organic ‘litter’ as well, which provides food for resident invertebrates. Others utilize their roots to help retain organic matter. Not suprisingly, many plants in this group provide ‘homes’ for many other animal species, generally invertebrates, within their ‘cups’ forming ‘local’ biotic communities. Over 300 animal species are found no where else but on and in these epiphytic and lithophytic plants. These animals, in return, provide a source of more nitrogen for the plants. (If this seems odd to you know that in our own local regions several species of mosquito spend there larval stages in the water held in hollows and ‘cups’ amongst the branches and hollows of trees as revealed by canopy studies of trees like Coast Redwood and Douglas Fir, even Big Leaf Maple, some of which are very specific to these limited environments. Think of our ubiquitous Licorice Ferns and several species of Moss and Lichen. This group can also include higher plants including various angiosperms or flowering plants.)
In the epiphytic species, within the Tillandsiodeae sub-family, which include the ‘Spanish Moss’ of the American South, these Bromeliads lack the impounding leaves and cups and rely instead on ‘trichomes’, little scales or hair-like structures on the leaves surface, through which the plants take up the water. These modifications work together and are expressed to a greater or lesser degree in the many species members. Across the Venezuelan Highlands and the many wetter/warmer valleys and canyons of the northern Andes, in ‘cloud forests’ of Central America, and as far north as Virginia in our South, many of these epiphytic Bromeliads can be found where frequent rains and/or condensate from clouds and/or fog, regularly replenish their needed moisture.
Other Bromeliads took a different pathway and have become very successful in arid, even desert, landscapes. Some of these, like the Puya spp., populate arid western regions of the same Andes Mountains up into southern Central America. These are terrestrial plants, relying on their roots to find the moisture that they need. Possessing a very similar rosette structure, often including their impounding leaves, these plants funnel the infrequent, and seasonal, precipitation they get into their crowns and root structures where they can take it up, a strategy very similar to Agave and Aloe which grow under similar conditions.
Over the course of millions of years, as the highlands pushed up, as the Andes formed, Bromeliads adapted to the higher, drier, climates that had come available transforming their metabolism to what we now identify as the CAM photosynthetic pathway. These plants utilize solar energy captured in the daylight hours, transforming it into ‘ATP’, a metabolite that provides ready energy for other chemical reactions, working as a kind of biologic ‘battery’ that can be charged, spent and recharged, as do all photosynthetic plants, but this time producing the organic compound, crassulacean acid, ‘storing it’ until nightfall, when leaf stomata open allowing in the CO2, then completing the process transforming it into the carbohydrates and manufacturing the other metabolites that the plant later uses to power its cell growth and build its needed tissues and structures. In CAM plants these two stages are separated over time effectively and greatly lowering water loss by the plant, minimizing the effects of high ambient temperatures, but, in the process, greatly slowing the overall growth of the plant. C3 plants (Worldwide the most commonly utilized ‘pathway’, produces a 3 Carbon molecule as an intermediate step instead of Crassulacean Acid, or the 4 Carbon molecule utilized in the intermediate C4 process.) grow more rapidly, wasting considerable water in the process as well as energy through the less controlled respiration, or oxidation, that can occur within the plant when the processes can occur simultaneously, breaking down some of the carbohydrates, that the plant, has previously produced. C4 plants, thought also to be an adaptation to drier conditions, separate the internal chemical processes physically in such a away that, oxidation, respiration losses, are minimized and are most commonly found in dryland grasses. CAM is the most water efficient pathway. (See previous postings for a more detailed description of these alternate forms of plant metabolism.)
Succulents or not? and Drought Response



Having described Bromeliads as water thrifty plants that utilize the CAM photosynthetic pathway, I need to say that this does not make them succulents. Most succulent plants aren’t epiphytic. Some are lithophytic like the Dudleya that populate some of our west coast rocky points and cliffs. I’m not sure if Dudleya have the structures or ability to take moisture from the air or if they, like terrestrial plants, depend on their roots for water uptake. Succulents come from low water areas, often desiccating, environments and, like Bromeliads, utilize the CAM pathway. Succulents also utilize other strategies to address drought with their capacity to store water within their tissues, a thick cuticle and or other adaptation to their epidermis to reduce water loss and having shallow root systems able to absorb water from even light, non-drenching showers. All plants contain water within their cells, tissues and, in those that have them, being moved through themselves ‘freely’ within their vascular systems. Plants that are considered succulents have the capacity to ‘store’ water for later use when it becomes too limited in their surrounding soil. This water tends to be ‘bound’ within the plant in the form of ‘gels’ which is water bound to complex proteins in the plant and can hold a great deal of water until conditions change within and a signal is sent to release it. This is not simply an on/off switch within the plant. The need is very ‘localized’ within the plant. Many plants commonly recognized as succulents occur within the Crassula (this is the group of plants within which researchers first identified the CAM pathway), Cactus, Agave and Euphorbia families, among many others. They occasionally occur as woody plants like the Australian Bottle Tree, Brachychiton rupestris and the genus Pachypodium, but none of the Bromeliads are succulents.
Succulents have thickened leaves and/or stem tissues, some include root structures as well, that can hold water that is not being actively utilized metabolically to enable and support growth, nor is it being moved directly to cells where it can be ‘immediately’ used in this way. During low water periods it can be moved internally to where it is needed resulting in a shrinking or withering of these leaf and stem tissues, as internal organic structures, likely proteins, release it, when the drought period is long enough. In wilting, plant cells and tissues, begin to dry.
[Some species, typically those C3 plants coming from wet climates, can wilt in response to heat if it is too high for them to carry on with their growth processes. Their metabolic processes can be shut down directly because of ambient temperatures that are too high. This begins to happen in most plants by the time 90ºF is reached. The addition of water to such plants will not help them and may in fact lead to their rotting and the promotion of fungal diseases. What these plants need are lower temperatures.]
Under-hydrated plants will begin to wilt if they have leaf structures subject to it. Many succulents don’t have leaves, or have leaves that are extremely modified, minimizing their water loss from their surfaces, while in others their leaves contain water storage tissue. Water serves a structural function within the cell as well as its vital metabolic functions. Any plant can tolerate water ‘scarcity’ only to a point. If it goes too far or extends too long, the tissues ‘permanently wilt’ and its cells are physically or structurally damaged. These will not revive with the addition of water. When this point is reached, cell structure collapses, and the cell dies. Succulents can also wilt and die in extended drought, but their capacity for it is much longer due to their water storage ability…and their ability to call on those reserves. Some deciduous plants have the ability to enter drought dormancy and they do this in part by purposely dropping their leaves. They are genetically adapted to this and are not unduly stressed by it. Plants that don’t possess this capacity will attempt to regrow/replace all of the leaves and other desiccated tissues it has suffered, leading to more stress, weakening it going into its winter/cold dormant season. For many plants this forced defoliation may be enough to kill the plant out right.

This was my Agave ‘Sharkskin’ last summer. A monocarpic succulent, Agave draw on their moisture stores when they literally erupt into flowering, drawing moisture from their leaves while utilizing their stores of carbohydrate. While this plant is ‘not’ drought stressed you can see what happens to the leaves as they lose moisture shrinking and distorting them.
Succulents are able to withstand extended dry periods and recover without significant tissue loss unless the drought period is beyond its capacity. When drought stressed these plants begin to draw on their reserves, as long as they are available. They still require water. Drought tolerant does not mean a plant doesn’t need water, only that it can operate with it at relatively infrequent or low levels of availability, as long as it is available in adequate amounts so that it can replenish itself…like any plant. Drought tolerance is the ability of a plant to ‘bridge’ the periods of low water availability a particular species has adapted too. One plant, succulent or not, may be ‘drought tolerant’ in one setting, but not in another drier setting. Some Bromeliads like Puya and Hechtia can be extremely drought tolerant because of their adaptations….They just aren’t succulents.
Bromeliads’ above ground structures are primarily leaves. With a few exceptions they have no significant stem tissues to ‘hold’ water even if they could. Their leaves are relatively thin in section and tend to be more ‘rigid’ then those of many plants. They regulate their water use metabolically by utilizing the CAM pathway. They cannot ‘swell’ with ‘extra’ water, forming storage ‘gels’ within their tissues, nor ‘shrink’ when it is drawn away for plant use. All of the water within the plant, intra- and extra-cellular, is necessary for the healthy function of the plant. Rigid leaf structures are often found on drought and very cold tolerant plants, like the sclerophyll native to chaparral and some steppe regions, but not always. As a group the family is intolerant of significant or sustained freezing temperatures. Cold hardiness depends on more than a plant having rigid tissues…but, many terrestrial Bromeliads are quite drought tolerant relying on their water ‘frugality’, their significant and active root systems and, in some cases, the thick, tough epidermis of arid terrestrial species. These possess the scale like, trichome structures, that in their case, protect them from the burning and drying of direct and intense sunlight. Their rigidity does help provide the structural integrity they need to both ‘defend’ themselves as I discuss elsewhere and, for those that are epiphytic and have impounding leaves, aid their ‘ability’ to hold water within the cups formed by their rosettes.
Puya



The name “Puya” comes from the Mapuche Indian word meaning “point”…the assignation is clear and the pointed, spiky, nature of this genus is immediately obvious to anyone. But there is something easy and comfortable about the sound of the word in your mouth when you speak it…poo-‘yah. Puya are native to the arid portions of the Andes and South American western coastal mountains. (Oddly, two species are found in dry areas of Costa Rica.)
My Puya chilensis, is native to the arid coastal mountains of Chile where they share space with other sclerophyllous plants, all characterized by their small, evergreen, stiff leaves. Interestingly it can be found growing near the endangered palm, Jubea chilense and a bamboo, Chusquea cumingii. This area is sandwiched between the Atacama Desert to its north and the Lakes District to its south. The Atacama, considered to be the driest desert in the world, stretches a 1,000km north and south abutting the western flank of the Andes, in its rain shadow, a result of the predominant east to west winds at these latitudes. In Chile, these landscapes are called mattoral and are very similar to California’s chaparral and share their ‘mediterranean’ climate. This Puya is most likely to be found on north facing hillsides, between 1,000′ and 3,200′. Some of these large Puya species are considered protocarnivourous, entangling animals that are thought to ‘feed’ the soil and then the plants, as they decompose. I can almost imagine their terror at becoming entangled with each struggling move only worsening their plight! This is another acquisition for me from Annie’s Annuals, I couldn’t resist her description…we shall see how long I can keep it! It forms a rosette 5’ tall with truly formidable foliage as you might have already guessed. The flowers though are the aesthetic ‘hook’ forming on 12’ stalks and blooming for a few months beginning in Spring. It takes 10-15 years to bloom, sometimes as long as 20, but than blooms every year after…one of the few Puya that are not monocarpic.
Puya chilensis

My Puya chilensis, small and mean at this stage. If my tape measure looks upside down, recall that this is a southern hemisphere plant.
My P. chilensis is confined to a pot, which, because of its ultimate mature size will in all probability keep me from growing it into its flowering maturity…yet I have it! It will be big and heavy, two characteristics that will cause its lifting and handling ‘delicately, to be difficult, and, consequently, a more ‘bloody’ affair. This plant has to be a ‘9’ on the ferocity scale, it gains in fierceness, with increased stature. As terrestrials, Puya depend on their roots to take up moisture so unlike their epiphytic cousins and true succulents these will demand relatively large soil volumes/pots to grow into healthy specimen. Additionally, confined in too small of a pot many terrestrial Bromeliads are difficult to water, simply getting a hose or the spout of a watering can to where they can reach surface potting soil, can be difficult due to how the stiff downward arcing foliage can shed much of water out and away.
Puya raiimondii



Each of the around 200 species, have their own particular range and preferences. Puya raiimondii, Queen of the Andes, is restricted to the arid high Andes of Peru and Bolivia, where it can be found in the Altiplano, an arid high elevation plateau, averaging over 12,000′, lying between two ranges where the Andes are at their widest. The ‘high plain’ stretches south from central Peru across Bolivia, where Lake Titicaca is located near their shared border, all of the way down to northern Argentina and Chile, the bulk of it in Bolivia. It is bound to the east by the Amazon Basin across more mountains and the Atacama Desert to its southwest. Active volcanos abound in the surrounding mountains. It is a huge basin out of which no rivers flow. The plateau averages less than 8″ of rain a year which it receives mostly in the summer months. The rest of the year is cool, dry, windy and sunny…desiccating conditions. Keep this in mind. You should consider this pattern for any of the Puya as most are from the western Andes growing under similar conditions.

Puya raimondii growing in situ on the Altiplano in Peru. credit to Pepe Rocque, Wikipedia
P. raiimondii, is found primarily, in the ‘warmer’ northern portion into Peru, including Huascarán National Park which covers over 1,300 sq.mi. It typically does not freeze where it grows, this is a ‘tropical’ area despite its elevation. It would then prefer to neither freeze nor bake. It is the largest and tallest of the genus and family, its vegetative rosette base reaching nearly 10’ tall, its inflorescence to 23’+ even up to 29′, carrying as many as 20,000 flowers, producing upwards of a million seeds. This species can take up to 40 years to bloom and, being monocarpic, like most Bromeliads, then dies. This particular species is considered endangered. It is unlikely that you will find one to grow on.
Puya mirabilis






Obviously then, all of the other species are smaller. Puya mirabilis, its rosette only 10”-12” tall, which I grow in a pot, is reportedly the quickest to bloom… mine has begun after having purchased it in a 4” pot in Oct. ’15. This plant appears more ‘lush’ somewhat softer and greener than its cousins above. The flowers are yet to emerge, so going by Annie’s Annuals description they are, “elegant, flaring, lime green bells held in fuzzy purple-grey calyxes”. Like all monocots their flowers are three petaled, though these are much larger and more prominent than others that I’ve seen and will be quite prominently displayed on 3’ stems. This too is a Bolivian species and is very drought tolerant, deer resistant and loved by hummingbirds when in bloom. Hardy down to 20ºF, mine will be forever in a pot which suits its stature well. Of all of the Puya I’ve grown and/or researched, this might be the ‘best’ one to grow for the mild temperate gardener: relatively quick to flower, pretty flowers, a very manageable plant for a pot, drought tolerant/mediterranean and perhaps the ‘kindest’, least aggressive foliage of the genus, I’ll give it a 2 on the scale.
Puya venusta



Puya venusta…this is a sad story. I bought this one from Sean Hogan that he’d identified as the ‘pink stemmed form’ on the label. This is a beautiful, almost white leaved plant, with stiffly arcing, channeled leaves with the typical recurved hooks you would expect running continuously along their margins…but they are in scale with the plant’s stature, of up to 40” tall the offsetting rosettes can spread to 5′ across. The flowers rise up about 3′ above the rosette. Native to arid mountainous areas of central Chile and Argentina were it grows on sites typically with a northerly aspect, generally out of full direct sun. It tends to inhabit areas that don’t freeze though it is thought to be safe into zn 9. Some growers, like Ian Barclay of Desert Northwest, write that they expect this to be hardy down into the low 20’sF. Annie’s says that it should be tough down to 20ºF. Some rate it as a zn9 plant. This selection was collected west of Tafi del Valle, in Argentina at 11,800’, Sean is generally very good with his collection data and this emboldened me…not always a good thing. After growing it for a few years in a pot that I would protect from freezing weather by shuttling it down to my lighted basement, I decided to try it outdoors, in the ground. I planted it in my south facing parking strip at the base of an established Trachycarpus fortuneii, 12’ or better tall, my thinking going somewhere along the lines that the tree would protect it from radiant heat loss and shed some of our winter precipitation away. It did this. The tree’s roots also helped keep the winter soil drier as well. This seemed to work well for its first two winters, both of them mild, but it was a mistake. This last winter was much wetter than is normal for us (See the comments under the chart above). Combine that with the two severe, for us, cold snaps, one in early December and one a month later, and the latter dropping to about 15ºF and tormenting us and our gardens with freezing and near freezing temperatures in early December and then again throughout much of January … this plant perished.
Puya dyckioides




Puya dyckioides, another Andean Puya from Bolivia and NW Argentinia, has been successful for me outdoors. It’s been growing on my south facing retaining wall, protected by an overhanging Oleander. It has formed a substantial clump, newer offsets crowding out older, blooming consistently over the last several years. It is relatively easy to work in and around as the leaves are neither overly stiff nor are they heavily ‘armed’. You still should be careful and deliberate around it, but the risk of injury on a scale from vicious and unavoidable, as a 10, down to ‘you have to try to inflict injury upon yourself’ as a 1, I’d put in the 2-3 range. The plant has an overall softer, ‘grass like’, appearance. The marginal spines are tiny. Still, while grooming it earlier this summer, reaching in and pulling away the older dead rosettes, my unprotected forearms received many tiny and irritating little jabs, that brought up no blood, but left my arm looking as if it were covered with tiny ‘hives’ and itchy. These take 6-7 years to reach blooming size and I believe this is my fourth summer with bloom. If your sun is too intense, as my site would be for this plant, offer it some shade.
Puya berteroniana
Puya berteroniana, possesses more than promise for our region, it has demonstrated some durability in the area already. I got two plants a couple of years ago from my friend Mike Remmick. Mike’s greenhouse is in the hills south of McMinnville, OR. I’ve come out and helped him a couple of times cleaning up and reorganizing it in late Spring and noticed his collection of Puya spp.. Mike has visited South America on plant trips and like’s this species…he also felt it should be tough enough for our typical winters, temperature wise, and experimented at his previous home in McMinnville planting it out in the garden…it survived and performed. He has since moved into SE Portland and Puya berteroniana has found itself out into the hell-strip.



I’m lifting the following from Annie’s Annuals website…who wouldn’t want to grow this!
“One of the most astonishingly beautiful plants to grace our planet direct from Chile! Spectacular, 2-3’ long and 1’ across pineapple-y shaped spikes are densely studded with 1.5” waxy, metallic blooms of the most unearthly emerald-turquoise. Contrasted by bright orange stamens and filled with blue syrupy nectar, they’re a bonanza for birdies who perch on sterile, horizontal stems. To 6-10’ tall in bloom (year 6-8). Spreading slowly by offshoots, spiny 3’ silvery rosettes work well as a security barrier. Can also be grown in a container! Deer resistant, plant in well-drained soil with occasional Summer water. Hardy to 20°F.
Check out the description on the Strange and Wonderful Things site, the creators of which wax on even more eloquently also giving me some question as to where to plant this as over time they can offset and form extensive colonies…a problem that I don’t anticipate here!

Growing in situ. Image of Puya berteroniana VII Region, Camino a Laguna Maule, Chile Altitude: 500-2200 m. 01 13, 2006
This Puya is endemic to the mediterranean climate of the coastal and Andean mountains of central Chile, between 500m (1,600′) and 2,000m (6,500′) where it experiences a dry summer period, much like ours, of 3-5 months, but can go much drier tolerating as much as 10 months of drought. It’s precipitation ranges from as little as 4″ annually, but more commonly between 16″- 32″. Chilean experts rate it hardy to -5ºC (23ºF), but again, Mike has grown it successfully in McMinnville, USDA 8a, 20ºF. I did find other sites that rated it down to 15ºF! Most Puya are thought to be intolerant of freezing temperatures. This is a full sun plant and is typically found on sloping sites, often with coarse mineral soils.
I’ve grown my two plants into crowded 2gal size, one of which I planted on my south facing retaining wall, this last Fall, in a spot where I’d just removed a Brown Turkey Fig tree that had become an out of scale and almost unharvestable beast even with my heavy pruning of it. The new Puya shares the bed with other plants like the blue form of the Mediterranean Fan Palm, an Oleander, prostrate Rosemary and my well established Puya dickyoides, described above. It will get a little shade here from a Chinese Fan Palm growing to the south of it…but I’m hoping it will do okay. As noted above these take 6-8 years to bloom, but then become perennial bloomers…this is no one and done performer.
Before I leave this species, I most mention that yes, it does register well into the danger region on the ferocity scale, being ever so slightly less fearsome than Puya chilensis, I’ll give it an 8+…so, if you choose to try this, be forewarned!



I am tempted to try P. venusta again and am intrigued by P. alpestris, they aren’t too huge, and can take some minor freezing, so I won’t need dedicated full time indoor space for them all winter, so for the most part they can stay out on the covered porch dry. These species feature, colorful and spiky inflorescences….and yes, those aggressive recurved marginal spines as well.
The Fascicularia and Ochagavia Question

The inflorescence of Fascicularia bicolor with its blue flowers. The center of the foliage on a blooming rosette, turns red when the flowers appear and then, like many Bromeliad, that rosette dies replaced be previously formed offsets. My plant has yet to bloom so this photo comes from Wikipedia.
If you’re not into blood and guts, consider this genus, as on my scale of one to ten as described above with ten warning of near complete evisceration if one is fool hardy or reckless, this one’s a solid 4, dangerous enough but not stupidly so. Fascicularia pitcairniifolia. You would think that in a genus composed of one or two species things would be pretty well settled taxonomically, guess again. Originally described as F. bicolor it was reclassified as F. pitcairnifolia and later changed back to F. bicolor. Subspecies were proposed. Changes retracted. There are significant differences in the sampled populations, but were they sufficient to constitute separate species??? Adding confusion at a different level are those who say the species name indicates that it is from Pitcairn Island. It is not. The specific epithet simply recognizes a similarity to the foliage in genus, Pitcairnia, another Bromeliad member. This Fascicularia is from the lower Chilean Andes, allegedly north of the other Fascicuaria species, F. bicolor which is suppose to be slightly hardier and occurs at least occasionally as an epiphyte! Some botanists have argued that F. pitcairnifolia possesses thicker, slightly wider leaves. and some minor differences in the timing of flowering and is reputedly slightly less hardy. The ranges of both overlap Good luck sorting this out.
They say that there is no shame in trying, that the shame is in avoiding the challenge…don’t they say this? Well, I bought this one from Sean, in 2012. Grew it in a pot for awhile and then planted it out in ‘13 on my east facing retaining wall, a rotting old RxR tie affair. My plant failed to grow the following spring. [trans. it died.] It’s demise is recorded in my database, winter of ’14. What happened to it?
That winter our temperatures dropped five times down into freezing temperatures, each time for several days in row. Over two of those periods, one in early December the airport dropped to 12º and another in early February, when we dropped down to about 20º. At the same time we recorded sub-freezing highs with our usual pattern of at least several days above normal high temperatures in-between, before freezing, even lightly again. In other words, the typical yo-yoing that we get that can wreak havoc with a plant’s dormancy making them more subject to damage or death. The rain year measured from Oct. 1 to Sept. 30 was 5” below normal. So, what killed it? Some growers in England say that with excellent drainage, these can go as low as 14ºF. Well, mine was planted in the local loam right next to the wall, but it was likely still too wet. Wet or cold? They tend to compound the negative effects of each other. I won’t be trying that again without a backup plant in a pot and I won’t try it unless I’m planting into scree or a well constructed crevice bed.


Fascicularia bicolor ssp. caniculata ‘Spinners’. I bought another one of these from Sean out a Cistus in June of ’14. I potted it up a couple times in consecutive years leading into the summer of ’17. I plant it in my typical mix with added pumice making it nearly half pumice. Once every couple weeks I set it in a tray to bottom water it along with a ‘squirt’ to the top. Sometimes I’ll give it an additional squirt between these more thorough soakings. This is the schedule I follow with many of my potted succulents, though I avoid the top watering of those that the practice encourages fungal growth on. Watch and learn. This seems to work well for my more ‘arid’ terrestrial Bromeliads.

Zona Sur, or the Lake District, is depicted in light green on the map. 45º latitude, comparable to Portland, OR is just south of its southern border. Doing the ‘hemispheric flip’ would place Zona Sur along the central Oregon coast down into the northern California coast. The weather patterns and coastal currents are very similar, but the differences in land masses, and mountains result in different growing regions. The west coast of Chile of Zona Sur and on south is characterized by hundreds of islands, bays, inlets and fjords. The Zona Australe shares much with the coast of British Columbia.
Fascicularia bicolor grows primarily in scree, but can be found as an epiphyte in Chile south of Santiago in the Lake District, or Zona Sur. Chile, a very long narrow country, is divided into 5 different biological regions north to south with widely varying climates, the huge Atacama Desert, considered the driest desert in the world, stretching 600 miles from Peru along Chile’s northern boundary, south, lying within the area colored ‘coral’ to the left. Zona Sur lies between Central Chile, or Zona Central, the native range of several of the dryland Puya spp., defined by the Rio Bio Bio. Zona Australe is to its south, which includes the remote and cold southlands of Patagonia, the division being at the Chaco Channel; the Pacific is the obvious western boundary and the Andes and Argentina are to the east. The Zona Sur and Australe are deeply scarred by glaciation and can be characterized by abundant rivers and lakes sliced through by channels and fingers of saltwater. There is a Central Valley that slopes southerly down from the north into an inland sea, the Golfo de Ancud. Geologically it is a volcanic region with well drained soils and scattered younger peaks standing well above the eroded ridges and mountains. The climate is Valdivian Temperate Rainforest, extending well easterly into the mountains with rainfall totals from 100″- 200″. As you might guess temperatures tend to be moderate/cool. It lies between 37º S and 43.4ºS latitude and has strong seasonal westerly winds, not unlike our condition here along the Oregon and Washington coast. This forested country is dominated by southern hemisphere conifers and both evergreen and deciduous Nothofagus trees along with Myrtles. There are forests of Aurucaria and Fitzroya as well as large wetlands. Unlike the Puya, Fascicularia spp. are not arid region plants. They can often be found growing on trees or cliff faces where water doesn’t collect, draining and percolating quickly away. It is humid country. Some growers have recommended keeping it out of direct or intense sun which is probably more important if your summer temps trend above 85ºF. I move my pots around and none of them get full afternoon sun though they can get a couple hours at mid-day. With free draining or scree I might be tempted to grow this in the ground, I might even try it in a shadier portion of my front retaining wall once it bulks up. Overhead protection, canopy, could be of help as well.

To better understand the scale of a place like Chile I include this. Chile is 2,356 miles long north to south and averages 177 miles east to west. Here the mapmakers have rotated Chile 90º overlaying in on the US. Both are drawn to the same scale. Zona Sur, on this map, would cover Chile from central ‘Wyoming’ to central ‘South Dakota’. Santiago, the capital in central Chile, is at 33ºS latitude which would put it just south of Los Angeles in the Northern Hemisphere. Were Chile stretched out along the coast of North America it would reach from about 18ºN to 55ºN, roughly from almost Acapulco, Mexico, almost to Ketchikan, Alaska. Spanning such a distance north and south suggests how diverse its climates are. Add to this the ruggedness of the country, and the Andes with an average elevation of 13,000′ and you get an even clear picture of its differences. It is also interesting to note that Chile contains some 5,100 different species of plant almost half of which are endemic, occurring no place else on earth.
My friend, Mike, has been having some success growing this in the ground here in SE Portland where it is planted on a slight north slope at the edge of an old large, topiary chicken! Yes, a chicken…of the Boxwood type. It has been there only a year (spring ’19), so the verdict is not yet completely in. It would seem that as these mature and produce offsets the bulking up aids a bit in their hardiness and tolerance. Some others have suggested to me that these are somewhat temperamental and don’t divide them….I think that difficulties with division might have more to do with the timing and after care. When I divided mine each plant had very few roots. I would suggest doing what I didn’t do…I tore my entire plant apart rather than just a few offsets and the old spent rosettes…the result of which will be that it will be likely two years or so before I see flowering again. Didn’t really think that one through. I pin these in place to stabilize them as I’ve done when dividing other Bromeliaceae. For a simple how to and confidence builder, check out this video produced by Burnacose Nurseriers in the UK. Of course the larger the plant you are dividing, the more rosettes it has, makes this more of a physical challenge in addition to the issue of their lightly barbed leaf margins, perfect for tearing skin, which she doesn’t mention. They suggest doing so in winter but they have heated glasshouses and I do not. My basement can get quite cool in winter, down to 55ºF which will slow rooting.
One reason for dividing them, beyond their getting large and congested, is that over the years the rosettes begin to flower, which is what we want, but the old inflorescences remain attached turning brown quickly, tightly nested in the base of the old rosette while it slowly declines. Like many Bromeliads this is monocarpic so the result will be old browning rosettes amongst your fresh, un-flowered, green ones. In a small pot or when viewing up close, this is an aesthetic negative, so I chose to divide, this time not doing so until the early summer of ’18. I wanted time for the plants to be able to anchor themselves before going into winter when moving the pots around you could disturb the slow growing roots.

Ochagavia carnea and Fascicularia inflorescences are very similar, which is consistent with their very similar genetics arising within the same ‘clade’ or genetic line….Individual flowers are around 3″ long and are packed tightly into a head, the inflorescence, which can be around 4″ across and tall.
A quick overview: the Bromeliad family is limited to the western hemisphere, the bulk of member species native to South America. There are 23 species of Bromeliaceae in Chile, twenty of which are endemic. The three species of Tillandsia found in Chile are not endemic. The local genus includes T. usneoides, Spanish Moss occurs commonly across much of the wider range of Bromeliads into North America. Fascicularia spp. and the Ochagavia spp. have very limited ranges and are endemic to their regions discussed here in Chile. Ochagavia include four species. O. andina, which as it’s name suggests, is a resident of the Andes, limited to a small area in Central Chile, in lowlands and valleys, from 2,300′ up to 8,000′ or the limit of occasional snowfall, well away from the coast. It is thought to be hardy to brief freezes of 23ºF, though relatively little is known of it as it is relatively rarely observed in the field O. litoralis, as it’s specific epithet suggests, is limited to the mild coastal region of central Chile where it grows above the heavy salt spray to 800′, saxicolous, on rock. O. lindleyana…I couldn’t find much on this species and, some don’t recognize it, would seem to have a very limited range, to below the line of snowfall in the Valpariso region, just north of Santiago. Other sources recognize O. elegans which has the most geographically limited range occurring only on the cliffs of Isla Robinson Crusoe of Juan Fernandez Archipelago where it is quite common, forming dense stands by offsetting freely.
I helped another friend divide a Bromeliad, I was told was an Ochagavia carnea, she had on ‘loan’ so three of us could each have a healthy sized piece. She hadn’t divided any of the Bromeliads before, I think also that she wanted to avoid any confrontations with the spiny margins. So I did it, happy to then have a plant of my own. Later I was told, by a knowledgeable third party, that this was in fact another form of Fascicularia bicolor!!! Apparently there is quite a range in the width and length of the leaf blades, as to my eye the plants looked significantly different. Ochagavia carnea has strappy leaves that can grow up to a meter in length, but can be, I found, half that. My plant currently needs up potting as it is very crowded, its leaves are limited to about 18″….The article, “Revision of the genus Ochagavia (Bromeliaceae, Bromelioideae)”, Georg Zizka, Katja Trumpler, and Otto Zöllner published by the Botanic Garden and Botanical Museum, Berlin, acknowledges the confusion both between and within these two genera. It would seem that there is work yet to do by the botanical specialists!…The only thing left for me to do is to grow them both out to blooming and then make a determination!!! Okay!

From the UBC Biology 343 Blog. The epigynous tube is the style with the stigma atop it. In Ochagavia spp. the center tube is as much as three times as long as the stamen, the reverse of the relationship shown above.

Ochagavia carnea individual flower. In the drawing (a) you can see the defining, exserted, style and stigma rising above the stamen. Taken from the above referenced paper from Botanical Garden and Museum Berlin.
Their inflorescences are similar in structure but the base of the leaves of the flowering rosette on O. c. turn pinkish, rather than the vibrant crimson red hue of the Fascicularia in question, with much less of the leave’s bases changing color. What botanists use to define the four Ochagavia spp. apart from each other most accurately are the comparative lengths of the stamen and pistil structures, notably, their prominent epigynous tube, as there is considerable overlap of their other structures. On Ochagavia spp. the epigynous tube can be as long as .75″, as much three times the length of the surrounding stamen. The Fascicularia bicolor and Ochagavia carnea have somewhat different flowers, blue verses pink, with slightly different sized petals, overall, their petals separate, overlapping at the base, with all of their parts in multiples of three, consistent with the flowers of all Monocots. Shouldn’t I be able to tell them apart? It is also interesting to note that the finer textured F.b. ‘Spinners Form’ flowered within 3 years of my purchasing it, bulking up quickly over the period. The coarser/larger O.carnea I have has bulked up quickly as well, though not to its described maximum mature size and has not flowered. I should note here that gardening experience in southern Europe, even on the island of Siciley, where O. carnea has naturalized to an extent, have been observed to rarely flower! Time to move on.
O. carnea is found frequently across its range within Central Chile, generally from 500′- 3,000′, away from the coast itself, but throughout the coastal mountains and on into the interior valleys of the Andes, occasionally higher up to the line of snowfall, north to the region of Valparaiso, above Santiago, and south to the Rio Bio Bio where the Zona Sur begins. It tends to occur as a component of sclerophyllous forest, comparable the California’s chaparral, or temperate deciduous forest in the southern portion of its range. They are most often found on rocky banks of streams and ravines, but can be found growing on a variety of sites from south facing, to flat, to north facing where they are protected from more intense sun. It would appear that this is the most moisture friendly of the genus. There is overlap of their range with several of the arid Puya species though the Puya are found on hotter more exposed sites. It is said to be tolerant of light freezes to -3ºC, but intolerant of snow accumulation. It is not found at all in the wet Valdivian Rainforest of the Zona Sur to the south. Across its range rainfall is in a mediterranean pattern and these are well adapted to 4-5 months of summertime drought, receiving between 15″- 31″ of rain over an average year. Some gardeners claim this is hardy to USDA zn9, or 20ºF, if its moisture and drainage requirements are met.
My plant spends most of our winters outside. I leave it on the porch where it can get rain, slide it under the roof if we’re going to have a light freeze and move it into the basement if its supposed to drop into the mid-20’s or lower. I’ll also move it under the roof when we’re in an extended period of heavy rain, keeping in mind that some years we can get nearly double their maximum rainfall these get in their native range.
Fascicularia is found more commonly in the cooler, wetter south of the Zona Sur, but the Fascicularia does range into at least the southern portion of the Central Zone where rainfall is lighter, so I can see how this physical overlap combined with morphological variations in these species can complicate field ID and a collector’s duty.
A brief update: we’re coming into spring of ’19 and the ‘Ochagavia’, or whatever it is, has increased in mass adding more rosettes to its bulk…but it still hasn’t flowered! I should pot it up and give the new mix a little slow release as the old soil is not doubt completely spent. Maybe it will bloom this summer, ’19???





















I live in Portland Oregon and my wife and I have had an old specimen of Ochagave carnea in our rock garden for years. We grow several Dyckia species and I am continually surprised by the hardiness of some of these terrestrial bromeliads. I’m growing a particularly rare Dyckia species- D. delicata from seed which I collected. When they grow to size, you would be welcome to have one if you are in the Portland area. My email is bachrocks10001@gmail.com
LikeLike
Anthony, my wife and I have been in and out of town over the last two months with more travel pending, for little vacations as well as dealing with family health issues so I didn’t contact you. I would love to come by sometime for an a visit of your garden sometime. We’re heading out again this weekend for a few days, but will be back by mid-week.
LikeLike